
¿Existen las especies transicionales?
Algunos negacionistas de la evolución esgrimen el argumento de que en la actualidad no existen especies "transicionales". Es decir, especies que hagan de nexo entre las especies actuales y sus precursores o ancestros.
Para ser rigurosos, cabe destacar que el término “transicional” viene del registro fósil: se habla de formas de transición o «fósiles transicionales» cuando un ejemplar muestra rasgos intermedios entre dos grandes grupos. Ahora bien, aquí conviene aclarar que es un término que hace alusión a un concepto "vivo" y relativo, ya que a decir verdad todos los seres vivos somos "transicionales", ya que no tenemos una forma "final" como especie. Esto es así porque la evolución no tiene una meta como tal, e incluso las especies actuales, con millones de años de evolución, no presentan una forma definitiva. La evolución es constante y continuamente desafía a lo ya establecido, generando mutaciones en los individuos que pueden desencadenar en una mejor adaptación al entorno en el que habiten.
Por lo tanto, y para que quede constancia antes de enfrascarnos en la lectura, decir que una especie “está a medio evolucionar” puede inducir a error: todas las especies vivientes están en algún punto del continuo proceso evolutivo (un proceso sin fin). Lo que sí podemos señalar son formas que conservan rasgos primitivos o que exhiben morfologías intermedias en el contexto de su linaje.
En muchos casos los ejemplos de transición (o “eslabones intermedios”) están en el registro fósil, no necesariamente en especies vivas que convivan con sus ancestros directos (porque normalmente esos ancestros ya están extintos debido a que evolucionaron en sus parientes contemporáneos, o bien fueron desplazados por otra especie).
*Durante la lectura encontraréis cruces delante de algunos nombres, indicando que actualmente dichas especies se encuentran extintas.
Ejemplos de formas intermedias o transicionales
A continuación se van a mostrar algunos ejemplos de formas transicionales que han existido o aun coexisten.
Tiktaalik roseae (final del Devónico)
- Nombre: Tiktaalik
- Especie: T. rosae (Daeschler, Shubin & Jenkins, 2006)
- Rango Temporal: 375 Ma
- Periodo: Devónico Superior (Frasniense)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata/Craniata
- Infrafilo: Gnathostomata
- Superclase: Osteichthyes
- Clase: Sarcopterygii
- Subclase: Tetrapodomorpha
Recreación del tiktaalik. Crédito: Tiktaalik roseae Daeschler, Shubin & Jenkins, 2006 http://spinops.blogspot.com/2015/03/tiktaalik-roseae.html

El Tiktaalik, un depredador de hasta 2,7 metros de largo, con dientes afilados, cabeza parecida a la de un cocodrilo y cuerpo aplanado, contaba con una anatomía y un modo de vida que se situaba a caballo entre los peces y los animales terrestres. Descrito por primera vez en 2006 y rápidamente apodado «fishapod», tenía características similares a las de los peces, como una mandíbula primitiva, aletas y escamas, así como un cráneo, cuello, costillas y partes de las extremidades similares a las de los tetrápodos (animales de cuatro patas).
El Tiktaalik era un pez sarcopterigio (aletas lobuladas) del periodo Devónico tardío que presenta multitud de características de los tetrápodos, considerándose un ejemplo de fósil transicional.
Vivió hace unos 375 millones de años. Los paleontólogos consideran que Tiktaalik representa un estadio intermedio entre peces como Panderichthys (que habitó hace unos 385 millones de años) y los primeros tetrápodos, como Acanthostega e Ichthyostega, que aparecieron alrededor de 20 millones de años después. La combinación de rasgos tanto de peces como de tetrápodos llevó a sus descubridores a describirlo como un “ictiotrápodo”. Tiktaalik se considera un fósil transicional tan relevante como Archaeopteryx. Entre sus características destacan:
- Branquias propias de peces.
- Escamas de pez.
- Extremidades con huesos y articulaciones intermedias entre pez y tetrápodo, incluyendo una muñeca funcional.
- Costillas robustas y un cuello móvil, propios de tetrápodos.
- Pulmones.
- Una región auditiva modificada.
Tiktaalik en el Field Museum, Chicago.

Modelo de Tiktaalik roseae en el Harvard Museum of Natural History


Eupodophis descouensi (Cretácico tardío)
- Nombre: Eupodophis
- Especie: E. descouensi (Rage & Escuillié, 2000)
- Rango Temporal: ~92 Ma
- Periodo: Cretácico Superior (Cenomaniense)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Tetrapoda
- Clase: Reptilia
- Orden: Squamata
- Suborden: Serpentes (basal)
Eupodophis descouensi es una serpiente fósil del Cretácico Superior (aprox. 92 Ma) descubierta en Líbano. Es uno de los taxones clave para comprender la transición evolutiva desde lagartos terrestres a serpientes modernas. Su característica más destacada es la presencia de extremidades posteriores reducidas, completas con fémur, tibia, peroné y metápodos, aunque sin función locomotora. Estas patas vestigiales apoyan fuertemente la hipótesis de un origen terrestre para las primeras serpientes, en contraste con teorías antiguas que proponían un origen marino.
Reconstrucción de Eupodophis descouensi (fíjate en el tamaño y la posición de las extremidades). Crédito: https://www.researchgate.net/publication/26386571_The_Cenomanian_Stage_of_hindlimbed_snakes
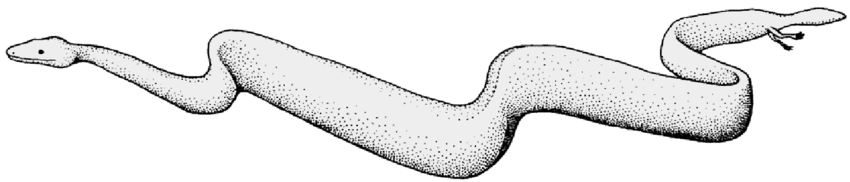
El esqueleto muestra un cuerpo elongado, numerosas vértebras presacrales y adaptaciones al reptado. La conservación excepcional del fósil ha permitido estudios detallados mediante tomografía de rayos X, revelando finamente la estructura de los huesos reducidos. Eupodophis constituye uno de los mejores ejemplos de estados intermedios (“formas transicionales”) en la evolución de los ofidios.
Eupodophis descouensi. Al Nammoura, Líbano, Cenomaniense. Regiones sacra y caudal y la pata trasera; la pata incluye el fémur, la tibia y el peroné, además de algunos huesos más distales. Crédito: https://www.researchgate.net/publication/26386571_The_Cenomanian_Stage_of_hindlimbed_snakes


Heteronectes chaneti (Eoceno)
- Nombre: Heteronectes
- Especie: H. chaneti (Davesne, Carnevale, Friedman & Barriel, 2012)
- Rango Temporal: ~50 Ma
- Periodo: Eoceno Inferior (Ypresiense)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Osteichthyes
- Clase: Actinopterygii
- Orden: Pleuronectiformes (forma basal)
- Familia: Incertae sedis (posición filogenética basal dentro del orden)
Heteronectes chaneti es un pez fósil del Eoceno temprano procedente de los famosos yacimientos de Monte Bolca (Italia). Es especialmente importante porque representa una forma transicional temprana dentro de los peces planos (Pleuronectiformes), el grupo que incluye lenguados, sollas y rodaballos.
A diferencia de los peces planos modernos —donde ambos ojos están en un mismo lado del cráneo— Heteronectes conserva aún una posición ocular casi simétrica, con un ojo ligeramente desplazado hacia el lado opuesto, pero sin haber completado todavía la migración craneal característica del grupo. Esto lo convierte en un excelente ejemplo de una etapa intermedia en el desarrollo de la asimetría cefálica.
Posee un cuerpo comprimido lateralmente, aletas bien desarrolladas y rasgos anatómicos que muestran la transición desde peces óseos simétricos hacia los altamente especializados peces planos modernos.
Una comparación de los peces planos vivos con otras especies vivas y extintas que dio lugar a este árbol evolutivo. Los «Pleuronectiformes» son las especies vivas de peces planos. b Una reconstrucción de Amphistium, el pariente más cercano de los peces planos vivos, muestra la posición intermedia de uno de sus ojos. c Una versión simplificada del árbol evolutivo de (a) ilustra la transición. A la derecha de cada pez hay tres vistas de su cráneo, con la posición de sus ojos marcada en rojo y azul. Crédito: Macmillan Publishers Ltd: Nature, 454 (7201):209–12, copyright 2008.
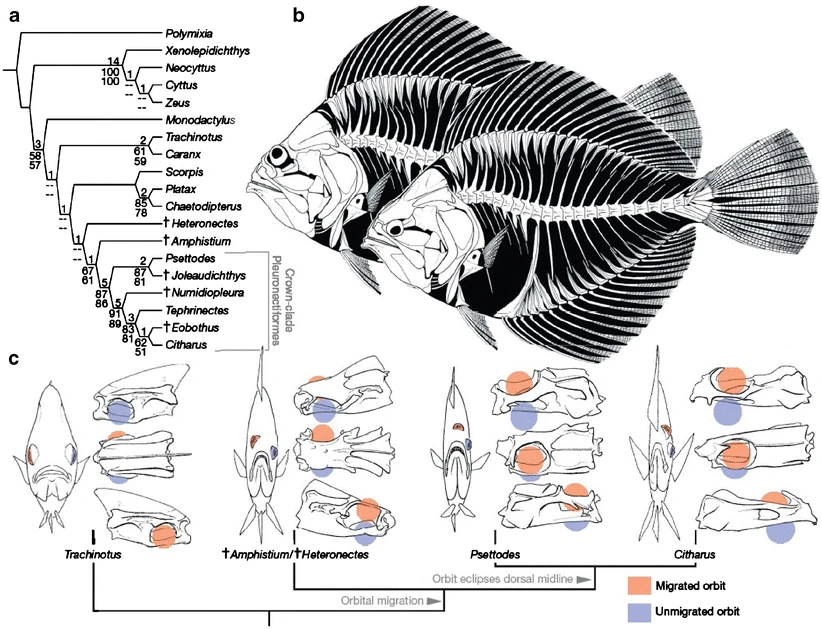
Amphistium, el pariente más cercano de los peces planos vivos, muestra la posición intermedia de uno de sus ojos.

Latimeria chalumnae (género de coelacantos vivos)
- Nombre: Latimeria
- Especie: L. chalumnae (Smith, 1939)
- Rango Temporal: Holoceno (linaje originado en el Devónico)
- Periodo: Actual
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Osteichthyes
- Clase: Sarcopterygii
- Subclase: Actinistia
- Orden: Coelacanthiformes
- Familia: Latimeriidae
Latimeria chalumnae, conocida como celacanto de Comoras, es uno de los casos más emblemáticos de “fósil viviente”. Se creyó extinta desde el Cretácico hasta su redescubrimiento en 1938. Es un sarcopterigio marino de gran tamaño (hasta ~2 m), nocturno y de movimientos lentos, que habita cuevas submarinas profundas en la región del océano Índico occidental.
Presenta aletas lobuladas y un esqueleto peculiar con un notocordio cartilaginoso, además de un cráneo dividido por una articulación intracraneal, rasgos propios de los sarcopterigios primitivos. Su anatomía y fisiología muestran un linaje que ha cambiado muy poco a lo largo de decenas de millones de años, siendo clave para comprender la evolución temprana de los vertebrados con aletas lobuladas.

Latimeria chalumnae. Crédito: https://www.fishi-pedia.com/wp-content/uploads/2023/05/coelocanthe-zoomcr.jp

Acanthostega gunnari (Devónico)
- Nombre: Acanthostega
- Especie: A. gunnari (Jarvik, 1952)
- Rango Temporal: ~365 Ma
- Periodo: Devónico Superior (Famenniense)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Osteichthyes
- Clase: Sarcopterygii
- Subclase: Tetrapodomorpha
- Orden: Ichthyostegalia (tradicional, para-etáxono)
- Familia: Acanthostegidae
Acanthostega gunnari es uno de los tetrapodomorfos más importantes del Devónico Superior, clave para comprender la transición del agua a la tierra. Procedente de Groenlandia (Formación de Ellesmere), vivió hace unos 365 millones de años. Representa una etapa temprana en la evolución de los tetrápodos, mostrando una combinación de rasgos acuáticos y características propias de los vertebrados terrestres.
Poseía ocho dedos en cada extremidad anterior, lo que demuestra que la pentadactilia típica de los tetrápodos modernos surgió después. Sus extremidades, aunque bien formadas, no eran capaces de soportar peso en tierra: estaban adaptadas principalmente para la propulsión subacuática. Conservaba branquias internas funcionales, cola con aleta y un cráneo aún muy propio de peces sarcopterigios. Estas características lo sitúan como un excelente ejemplo de forma transicional entre peces con aletas lobuladas y los primeros tetrápodos.


Archaeopteryx (transicional entre "reptil" y ave)
- Nombre: Archaeopteryx
- Especie tipo: A. lithographica (Meyer, 1861)(El género incluye varios ejemplares históricamente asignados a distintas especies, pero el taxón más común es A. lithographica.)
- Rango Temporal: ~150 Ma
- Periodo: Jurásico Superior (Titoniense)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Tetrapoda
- Clase: Aves (posición basal) / tradicionalmente Reptilia (Dinosauria)
- Clado: Dinosauria
- Subclado: Theropoda
- Infraorden: Paraves
- Familia: Archaeopterygidae
Archaeopteryx es uno de los fósiles más emblemáticos de la paleontología y un ejemplo clásico de forma transicional entre dinosaurios terópodos y aves modernas. Procede del Jurásico Superior (~150 Ma), conservado de manera excepcional en los depósitos litográficos de Solnhofen (Alemania), lo que permite observar detalles finísimos de sus plumas.
Presenta una combinación única de características:
- Rasgos avianos: plumas asimétricas desarrolladas para el vuelo, alas completas y un esbozo de furcula.
- Rasgos reptilianos/terópodos: dientes en la mandíbula, cola larga ósea con vértebras libres, garras en las manos y un esqueleto ligero pero no totalmente aviano.
Probablemente podía realizar vuelos cortos o planeos y se movía con agilidad entre la vegetación. Su mosaico anatómico lo sitúa como un punto clave en la evolución de las aves a partir de terópodos manirraptores.

Ambulocetus (transicional entre mamíferos cuadrúpedos terrestres y cetáceos)
- Nombre: Ambulocetus
- Especie: A. natans (Thewissen, Hussain & Arif, 1994)
- Rango Temporal: ~48–47 Ma
- Periodo: Eoceno Inferior
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Tetrapoda
- Clase: Mammalia
- Orden: Artiodactyla (Cetartiodactyla)
- Suborden: Cetacea (posición basal)
- Infraorden: Archaeoceti
- Familia: Ambulocetidae
Ambulocetus natans (“la ballena que camina y nada”) es uno de los fósiles más importantes para comprender la transición evolutiva desde mamíferos terrestres hacia los cetáceos. Vivió hace unos 48–47 millones de años durante el Eoceno temprano, en lo que hoy es Pakistán.
Poseía una anatomía claramente anfibia:
- Extremidades robustas capaces de soportar su peso en tierra.
- Pies y manos palmeados, adaptados para nadar mediante movimientos ondulatorios.
- Columna y pelvis que permiten un estilo natatorio similar al de los cocodrilos.
- Narinas en la punta del hocico, no desplazadas hacia la parte superior del cráneo como en los cetáceos posteriores.
Probablemente acechaba presas en aguas dulces o estuarios, y su estilo de vida recuerda a una mezcla entre nutria, cocodrilo y ballena primitiva. Es un eslabón crucial en la evolución de los cetáceos desde artiodáctilos terrestres hacia formas totalmente acuáticas como Basilosaurus.

Reconstrucción de Basilosaurus cetoides. Crédito: https://es.wikipedia.org/wiki/Basilosaurus#/media/Archivo:Basilosaurus_cetoides.png

Australopitecus (transicionales entre "simios" y humanos).
- Nombre: Australopithecus
Especies principales:- A. anamensis (4,2 Ma)
- A. afarensis (3,9–3,0 Ma)
- A. africanus (3,3–2,1 Ma)
- A. bahrelghazali (~3,5 Ma)
- A. garhi (~2,5 Ma)
- A. sediba (1,98 Ma, posición discutida)
- Rango Temporal: ~4,2–1,9 Ma
- Periodo: Plioceno – Pleistoceno temprano
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Tetrapoda
- Clase: Mammalia
- Orden: Primates
- Suborden: Haplorhini
- Infraorden: Simiiformes
- Parvorden: Catarrhini
- Superfamilia: Hominoidea
- Familia: Hominidae
- Subfamilia: Homininae
- Tribu: Hominini
- Género: Australopithecus
Australopithecus es un género de homininos que vivió en África entre 4,2 y 1,9 millones de años atrás. Representa un grupo clave en la evolución humana, al mostrar una mezcla de rasgos primitivos y derivados que iluminan la transición entre los primeros homininos bípedos y el género Homo.
Características principales:
- Bipedismo habitual, con pelvis y fémur adaptados a la marcha erguida.
- Brazos relativamente largos y manos aptas para trepar, indicando un estilo de vida parcialmente arborícola.
- Cráneo pequeño en comparación con humanos modernos (volumen endocraneal 350–550 cc).
- Dentición con caninos reducidos, molares grandes y esmalte grueso.
- Dimorfismo sexual marcado en varias especies.
Incluye especies célebres como A. afarensis (“Lucy”), A. africanus, A. anamensis y A. sediba (este último a veces asignado a Homo, pero frecuentemente mantenido en Australopithecus).


Sinornis santensis (Cretácico temprano)
- Nombre: Sinornis
- Especie: S. santensis (Sereno & Rao, 1992)
- Rango Temporal: ~120 Ma
- Periodo: Cretácico Inferior (Aptiense)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Tetrapoda
- Clase: Aves
- Clado: Enantiornithes
- Orden: Enantiornithiformes
- Familia: incertae sedis (dentro de Enantiornithes)
Sinornis santensis es un ave primitiva del Cretácico Inferior (~120 Ma) encontrada en la Formación Jiufotang, China. Pertenece al clado de las Enantiornithes, un grupo de aves tempranas muy diverso, pero completamente extinto.
Este taxón es importante porque representa un punto intermedio en la transición entre las aves basales (como Archaeopteryx) y las aves más avanzadas del Cretácico. Aunque conserva ciertos rasgos reptilianos —como garras en las alas y dentición— muestra varias características avianas modernas:
- Esternón osificado con quilla, lo que indica una musculatura de vuelo más desarrollada.
- Plumas bien formadas y alas funcionales.
- Adaptaciones arbóreas, incluyendo pies para agarrarse a ramas.
- Cola corta con pigóstilo, propia de aves más derivadas.
Su morfología sugiere un estilo de vida arborícola y un vuelo ágil, siendo un eslabón clave en la radiación temprana de las aves cretácicas.

Chlamydoselachus anguineus (Tiburón anguila)
- Nombre: Chlamydoselachus
- Especie: C. anguineus (Garman, 1884)
- Rango Temporal del linaje: Cretácico Superior – Actual
- Periodo: Holoceno (especie actual)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Deuterostomia
- Filo: Chordata
- Subfilo: Vertebrata / Craniata
- Infrafilo: Gnathostomata
- Superclase: Chondrichthyes
- Clase: Chondrichthyes
- Subclase: Elasmobranchii
- Orden: Hexanchiformes
- Familia: Chlamydoselachidae
- Género: Chlamydoselachus
Chlamydoselachus anguineus, conocido como tiburón anguila, es uno de los elasmobranquios más primitivos que aún existen. Aunque es una especie actual, representa una línea evolutiva muy antigua, con fósiles del grupo que se remontan al Cretácico Superior (≈80 Ma). Su aspecto serpentiforme y ciertas características anatómicas le han valido el apodo de “fósil viviente”.
Sus características más distintivas incluyen:
- Cuerpo alargado y flexible, similar a una anguila.
- Seis pares de hendiduras branquiales, con los bordes anteriores extendidos y unidos, formando una especie de collar (de ahí Chlamydoselachus: “tiburón con branquias frilladas”).
- Mandíbulas altamente extensibles, con dientes tricúspides curvados hacia dentro, especializados en capturar calamares y peces de aguas profundas.
- Aletas dorsales y pélvicas retrasadas, en una disposición primitiva.
- Hábitos bentopelágicos, viviendo entre 200 y 1500 m de profundidad.
Su combinación de rasgos antiguos y estilo de vida críptico lo convierte en un excelente representante de linajes basales dentro de los tiburones modernos.

Limulus polyphemus (cangrejo herradura)
- Nombre: Limulus
- Especie: L. polyphemus (Linnaeus, 1758)
- Rango Temporal del linaje: Ordovícico – Actual
- Periodo: Holoceno (especie actual)
- Dominio: Eukaryota
- Reino: Animalia
- Subreino: Eumetazoa
- Superfilo: Ecdysozoa
- Filo: Arthropoda
- Subfilo: Chelicerata
- Clase: Xiphosura
- Orden: Xiphosurida
- Familia: Limulidae
- Género: Limulus
Limulus polyphemus, conocido como cangrejo herradura del Atlántico, es un quelicerado marino cuya línea evolutiva se remonta a más de 450 millones de años, lo que lo convierte en uno de los ejemplos más emblemáticos de “fósil viviente”. A pesar de su nombre, no es un crustáceo, sino un pariente lejano de arañas y escorpiones.
Posee un cuerpo dividido en prosoma y opistosoma, protegido por un caparazón en forma de herradura, y un telson largo y rígido que utiliza para enderezarse si queda volteado. Dispone de cinco pares de patas, ojos compuestos laterales y varios ojos simples adicionales capaces de detectar cambios de luz.
Habita en las costas del Atlántico occidental y del Golfo de México, donde realiza desoves masivos en playas durante las mareas altas. Su sangre, rica en amebocitos, es extremadamente sensible a endotoxinas bacterianas y se utiliza en el test LAL (Limulus Amebocyte Lysate), fundamental para verificar la esterilidad de material médico.
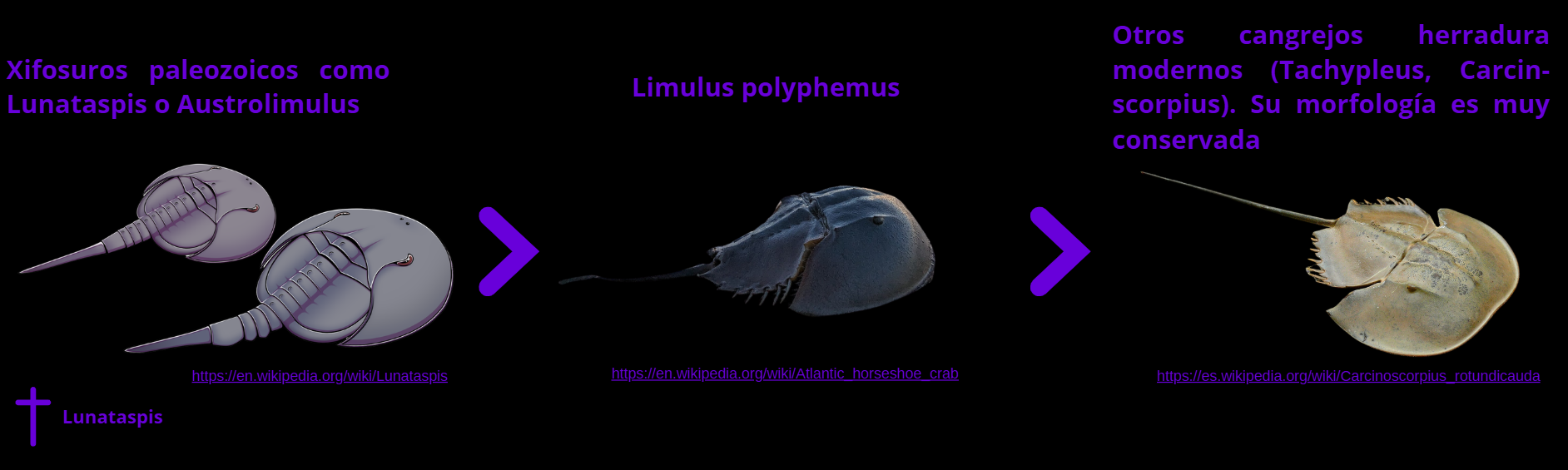
Fotografía de un cangrejo de herradura. Crédito: https://www.currentconservation.org/wp-content/uploads/2024/01/Copy-of-Horshoe-Crab-Tachypleus-gigas-in-Balasore-Photograph-by-Dr.-Biswajeet-Panda-scaled.jpg

Resumen de los ejemplos expuestos junto a otros de interés
| Nº | Especie / Género | Edad aproximada | Grupo de transición | Rasgos intermedios / significativos |
|---|---|---|---|---|
| 1 | Pikaia gracilens | ~505 Ma (Cambriano) | Cordados basales → vertebrados | Notocordio, músculos segmentados, cuerpo primitivo precursor de vertebrados. |
| 2 | Latimeria chalumnae (linaje ~400 Ma) | Actual, linaje muy antiguo | Peces de aletas lobuladas | Rasgos muy similares a los ancestros de los tetrápodos; ejemplo de morfología conservada. |
| 3 | Limulus polyphemus (linaje >400 Ma) | Actual | Artrópodos marinos | Morfología casi idéntica a fósiles paleozoicos; estasis evolutiva. |
| 4 | Tiktaalik roseae | ~375 Ma (Devónico) | Peces → tetrápodos | Aletas-limbs, cuello móvil, costillas fuertes: intermedio agua–tierra. |
| 5 | Acanthostega gunnari | ~365 Ma | Peces → tetrápodos | Dedos bien formados pero dependiente del medio acuático; branquias internas. |
| 6 | Ichthyostega | ~365 Ma | Peces → tetrápodos | Extremidades más robustas que Acanthostega, pero aún acuático; transición progresiva. |
| 7 | Ginkgo biloba (linaje ~270 Ma) | Actual | Gimnospermas antiguas | Morfología muy estable comparada con fósiles mesozoicos. |
| 8 | Eupodophis descouensi | ~95 Ma (Cretácico) | Lagartos → serpientes | Serpiente marina con patas traseras vestigiales funcionales. |
| 9 | Archaeopteryx lithographica | ~150–145 Ma | Dinosaurios → aves | Plumas modernas + cola ósea, dientes, garras en alas; forma transicional clásica. |
| 10 | Sinornis santensis | ~120 Ma | Aves primitivas → aves modernas | Esternón parcialmente desarrollado, muñeca similar a aves modernas. |
| 11 | Heteronectes chaneti | ~50 Ma (Eoceno) | Peces normales → peces planos | Asimetría craneal parcial: ojo a medio camino de migración. |
| 12 | Ambulocetus natans | ~48 Ma (Eoceno) | Mamíferos terrestres → cetáceos | Mamífero anfibio, locomoción mezcla de nado y marcha; oído semiadaptado al agua. |
| 13 | Chlamydoselachus anguineus (linaje ~80–100 Ma) | Actual | Tiburones antiguos | Mandíbula primitiva, cuerpo serpentiforme; morfología ancestral conservada. |
| 14 | Australopithecus (especialmente A. afarensis) | 3.9–2.9 Ma | Homininos tempranos → Homo | Bipedismo claro + cráneo y mandíbulas aún simiescas. |
| 15 | Homo erectus | 1.9 Ma – 100 ka | Humanidad temprana | Postcráneo moderno, cráneo aún arcaico; primera especie plenamente caminante y migratoria. |
Conclusión
Sí, existen formas con rasgos intermedios (“transicionales”) —especialmente en el registro fósil— que muestran pasos evolutivos entre grupos mayores. Pero conviene matizar que no hay especies “a medio evolucionar”, ya que todas las especies vivientes están en evolución y no hay jerarquía “más evolucionada” vs “menos evolucionada” en un sentido lineal.





{kind=link}
{kind=link}
{kind=link}