
La hipótesis de la Reina Roja

El nombre “Reina Roja” viene de un pasaje del libro A través del espejo de Lewis Carroll, donde la Reina le dice a Alicia:

“Aquí, como ves, hace falta correr todo lo que puedas para permanecer en el mismo sitio”.
En biología evolutiva, esta metáfora fue adoptada por el biólogo Leigh Van Valen en 1973 para expresar que las especies están en una carrera evolutiva constante: tienen que seguir adaptándose solo para “mantenerse en el mismo lugar” frente a otras especies que también evolucionan.
En otras palabras: una especie no evoluciona en un entorno estático, sino en un contexto donde sus competidores, depredadores, parásitos o mutualistas también están cambiando. Si no evoluciona, queda rezagada.
Van Valen usó esta idea originalmente para explicar por qué, en el registro fósil, las tasas de extinción no parecen disminuir significativamente con el “éxito” de una línea evolutiva: siempre hay presiones nuevas que empujan a las especies a cambiar.
Versiones de la hipótesis: macro y microevolutiva
La hipótesis de la Reina Roja ha evolucionado y diversificado según el nivel de análisis:
A escala macroevolutiva
Esta versión es más cercana al planteamiento original de Van Valen: trata de explicar patrones de extinción y especiación a lo largo de grandes escalas temporales, enfatizando que las interacciones bióticas (competencia, depredación, parasitismo) pueden ser tan importantes —o más— que los cambios abióticos (como el clima) como impulsores evolutivos.
En este contexto, la hipótesis sugiere que el “riesgo de extinción” para los linajes puede permanecer relativamente constante a través del tiempo porque el entorno biológico (los otros seres vivos) está siempre cambiando.
Una hipótesis alternativa con la que a menudo se contrasta es la hipótesis del bufón de la corte (Court Jester hypothesis), que sostiene que los cambios en el ambiente físico (clima, geología) son los motores más importantes de las dinámicas macroevolutivas.
A escala microevolutiva. Coevolución y reproducción sexual
En escalas más pequeñas (poblaciones, generación a generación), la hipótesis de la Reina Roja se aplica especialmente a las relaciones antagonistas, como huésped-parásito. El argumento es que los parásitos tienden a adaptarse rápidamente al genotipo más común del huésped, por lo que los huéspedes sexualmente reproductivos, al generar descendencia genéticamente diversa, tienen más probabilidades de “escapar” de infecciones especializadas.
Así, la hipótesis ha sido usada como una explicación del porqué la reproducción sexual es tan común (pese a sus costos) frente a la reproducción asexual: la recombinación genética ofrece una ventaja frente a los parásitos en una “carrera armamentista” evolutiva. Dependiendo de cómo sea la interacción genética entre especies antagonistas, pueden surgir distintos modos de dinámica (oscilatoria, escalada, persecución) dentro del marco de la Reina Roja. Algunos autores han descrito estas variantes como “Red Queen fluctuante”, “Red Queen de escalada” o “Chase Red Queen”.
Ejemplos y evidencia empírica
- En caracoles de agua dulce en Nueva Zelanda (Potamopyrgus antipodarum), se ha observado que la presencia de parásitos favorece comportamientos de apareamiento múltiples (lo cual aumenta la recombinación genética), como predice una versión de la hipótesis de la Reina Roja.
- En sistemas biológicos de virus o bacterias, se ha usado la idea de un “estado tipo Reina Roja” para describir cómo los patógenos siguen evolucionando para evadir el sistema inmune del huésped, pero sin que el patógeno “gane” del todo: debe seguir adaptándose.
- Modelos teóricos recientes abordan bajo qué condiciones la dinámica continua aparece en sistemas evolutivos como resultado de interacciones bióticas, es decir, cuándo una “carrera evolutiva perpetua” es robusta.
Tipos de dinámicas de Reina Roja
La “Reina Roja” no es un único proceso, sino un continuo de dinámicas coevolutivas, que varían según la naturaleza de la interacción (parasitismo, depredación, competencia, mutualismo) y el tipo de rasgos implicados (genéticos, cuantitativos o ecológicos).
Se suelen distinguir tres formas principales y una intermedia:
1. FRQ – Fluctuating Red Queen (Reina Roja fluctuante)
Definición:
La dinámica se basa en fluctuaciones cíclicas en las frecuencias de genotipos de huésped y parásito (u organismos en interacción).
Ningún bando gana de forma definitiva: cuando un genotipo se hace común, el otro evoluciona para explotarlo, y viceversa.
Características:
- Implica coevolución negativa (dependiente de la frecuencia).
- Basada en genes de efecto específico (matching alleles o gene-for-gene).
- Se mantiene la diversidad genética en ambas poblaciones.
- Típica de sistemas huésped–parásito o huésped–patógeno.
Ejemplos:
- Potamopyrgus antipodarum (caracol) ↔ trematodos (Microphallus spp.).
- Daphnia ↔ microparásitos.
- Peces espinosos (Gasterosteus) ↔ trematodos.
Resultado evolutivo:
Ciclos en las frecuencias de resistencia e infectividad → estabilidad dinámica sin tendencia direccional.
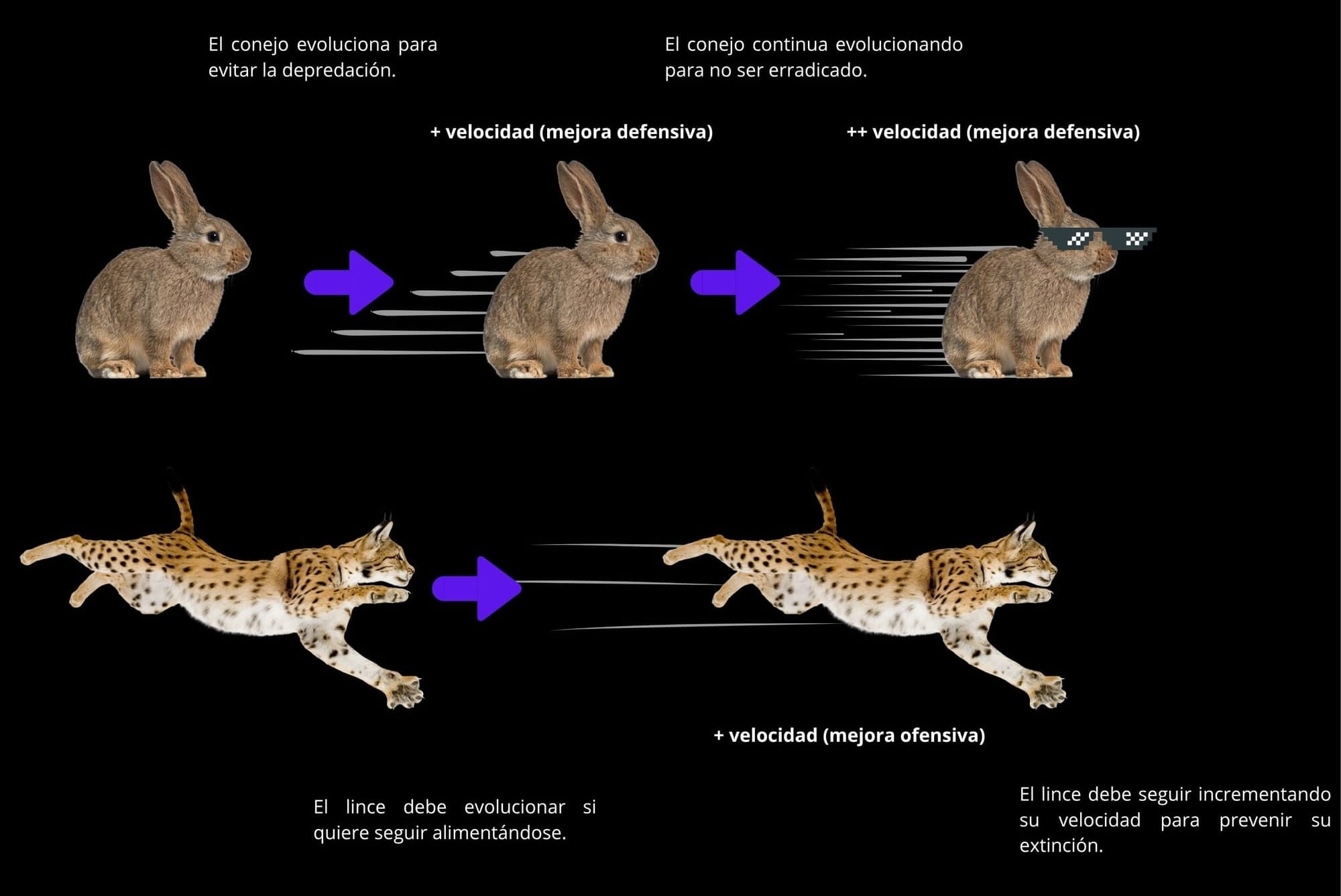
2. ERQ – Escalating Red Queen (Reina Roja en escalada o "arms race")
Definición:
Las poblaciones implicadas experimentan una escalada continua en los rasgos cuantitativos que determinan defensa y ataque: más toxinas, más resistencia, más dureza, más fuerza, etc.
Características:
- Implica selección direccional recíproca, no cíclica.
- Los rasgos evolucionan cuantitativamente (no por alelos discretos).
- Puede tener costes energéticos o fisiológicos crecientes.
- Típica de depredador–presa o planta–herbívoro.
Ejemplos:
- Tritones (Taricha) ↔ serpientes (Thamnophis): tetrodotoxina vs. resistencia nerviosa.
- Chirivía silvestre (Pastinaca sativa) ↔ orugas Depressaria: furanocumarinas vs. detoxificación.
- Camellia ↔ gorgojos perforadores: grosor del fruto vs. longitud del pico.
Resultado evolutivo:
Escalada de rasgos defensivos/ofensivos → aumento de complejidad o coste sin un vencedor final.
3. CRQ – Chase Red Queen (Reina Roja de persecución o ecológica)
Definición:
Cada especie responde adaptativamente a los cambios ecológicos o de nicho de la otra, no solo genéticos.
Una “persecución” donde los rasgos adaptativos cambian direccionalmente en el espacio o el tiempo.
Características:
- Afecta rasgos ecológicos o comportamentales (no solo genéticos).
- Puede implicar cambio de nicho, hábitat o estrategia trófica.
- Es más macroevolutiva: a escala de comunidades o especies.
- Común en coevolución espacial o “mosaicos coevolutivos”.
Ejemplo:
- Piquituertos (Loxia curvirostra) ↔ pinos Pinus contorta:
los pinos desarrollan piñas más grandes o duras; los piquituertos evolucionan picos más fuertes o curvados → persecución morfológica y geográfica.
Resultado evolutivo:
Cambio continuo del paisaje adaptativo → diversificación y especiación posible.
4. Mixtas o intermedias (FRQ/ERQ mixtas)
Definición:
Algunos sistemas muestran simultáneamente componentes fluctuantes y de escalada, dependiendo del gen o rasgo considerado.
Ejemplo:
- Linum marginale ↔ Melampsora lini (planta y hongo de la roya).
- Algunas interacciones gen a gen → FRQ.
- Otras (resistencia basal, virulencia) → ERQ.
- El resultado es un mosaico coevolutivo espacial y temporal.
Comparativa general
| Tipo | Mecanismo principal | Rasgos implicados | Dinámica temporal | Ejemplo típico | Resultado |
|---|---|---|---|---|---|
| FRQ | Dependencia negativa de la frecuencia | Genes específicos | Cíclica | Caracoles – trematodos | Mantenimiento de variabilidad genética |
| ERQ | Escalada direccional recíproca | Rasgos cuantitativos | Direccional | Tritones – serpientes | Arms race con costes crecientes |
| CRQ | Adaptación ecológica recíproca | Rasgos ecológicos | Espacial/direccional | Piquituertos – pinos | Cambio de nicho, diversificación |
| Mixta (FRQ/ERQ) | Combinación de ambos procesos | Múltiples tipos de rasgos | Mixta | Linum – Melampsora | Mosaico coevolutivo |
Importancia y límites de la hipótesis
Ventajas conceptuales
- La hipótesis de la Reina Roja desplaza el foco de la selección puramente ambiental hacia las interacciones biológicas como motor principal de cambio evolutivo.
- Proporciona un marco útil para pensar por qué muchas especies mantienen sexo, recombinación genética y diversidad interna, incluso cuando en apariencia podrían seguir un “camino óptimo” más sencillo.
- Ofrece una lente para interpretar cómo la coevolución puede generar patrones complejos como oscilaciones genéticas, cambios rápidos de estrategia, y estabilidad dinámica entre especies.
Críticas y límites
- En el registro fósil, no siempre se encuentran tasas de extinción constantes ni evidencia clara de que las interacciones biológicas dominen por completo las trayectorias evolutivas. Algunos cambios mayores podrían deberse a factores ambientales drásticos (clima, geología).
- No todas las relaciones entre especies son antagonistas; mutualismos y simbiosis también juegan roles importantes evolutivos. La hipótesis de la Reina Roja no necesariamente explica bien esos casos.
- Las condiciones para que la “carrera continua” se mantenga pueden ser bastante exigentes (por ejemplo, tasas de mutación, tamaño poblacional, costos adaptativos). Algunos modelos sugieren que en ciertos sistemas la evolución puede ralentizarse o detenerse — no toda población está en carrera perpetua.
Con este artículo damos inicio a la sección: Guerra biológica: carrera armamentística entre especies #1


Lecturas recomendadas (fuentes principales)


https://royalsocietypublishing.org/doi/10.1098/rspb.2014.1382

https://www.sciencedirect.com/topics/earth-and-planetary-sciences/red-queen-hypothesis





