

La Hipótesis de la Abuela: cuidar frente a parir. El enigma evolutivo de la menopausia.

La naturaleza es implacable. La evolución no negocia. Lo que funciona tiende a prevalecer, y lo que no, acaba desvaneciéndose. Sin más. Sin preguntar.
Todo parece responder a un balance coste-beneficio, a una economía elemental de la vida. Una característica que favorece la supervivencia o la reproducción puede abrirse paso generación tras generación; una adaptación cuyo coste no se vea compensado por algún beneficio acaba, tarde o temprano, perdiendo presencia. No siempre ocurre de forma limpia, directa, ni inmediata, pero esa tensión entre coste, ventaja y herencia ha modelado cada rasgo de los organismos que existen hoy en día.
Es el juego de la vida. Las reglas que nos han traído hasta aquí.
Y, sin embargo, a veces parece que los humanos vivimos al margen de ellas. Y es que, tras toda esta travesía evolutiva, podría parecer que hace tiempo dejamos de obedecer los criterios básicos que rigen al resto de seres vivos.
En efecto, el ser humano es una rareza. Muchas de sus adaptaciones son, como mínimo, desconcertantes: desarrollo tardío, infancia prolongada, piel frágil, escasa especialización anatómica, numerosos vestigios evolutivos, partos difíciles y crías extremadamente dependientes. A simple vista, no parecemos el diseño más eficiente de la naturaleza.
Y, aun así, hemos alterado el planeta como ninguna otra especie. Sin garras ni grandes colmillos, lentos campo a través y sin ser especialmente fuertes. En cambio, cooperación, lenguaje, técnica, memoria y transmisión cultural han formado parte de nuestra apuesta evolutiva. Nuestra ventaja ha residido en aprender juntos, acumular conocimiento y convertir cada generación en el punto de partida de la siguiente.
Quizá por eso algunas de nuestras rarezas no se entienden bien si miramos únicamente al individuo. En una especie tan social, tan lenta y tan dependiente del cuidado prolongado, la evolución no solo se juega en quien nace, sino también en quien consigue sobrevivir, aprender y reproducirse después.
Y entre todas esas rarezas humanas hay una especialmente intrigante: la menopausia.
Que una hembra deje de reproducirse no es un evento extraño. De hecho, tarde o temprano es lo normal. Eso, en sí mismo, no es lo extraordinario. Lo verdaderamente llamativo es lo que ocurre después, ya que en nuestra especie, muchas mujeres pueden vivir décadas más allá del final de su etapa fértil. Décadas en las que ya no tienen descendencia directa, pero siguen presentes, activas y profundamente integradas en la vida familiar y social.
¿Por qué? ¿Por qué la naturaleza apostaría esos recursos cuando la hembra ya ha cumplido con su compromiso vital?
Desde una lectura evolutiva superficial, esto parece una anomalía. Si la selección natural favorece aquello que contribuye a dejar más descendencia, ¿qué sentido tiene prolongar la vida mucho más allá de la capacidad reproductiva? Hay numerosas especies que mueren poco después de reproducirse, ¿por qué nosotros somos diferentes?
La respuesta quizá esté en que la reproducción humana nunca fue solo una cuestión de parir. A lo mejor también fue una cuestión de cuidar.
La contabilidad invisible del parentesco
Para entender lo que viene a continuación hay que cambiar la forma en la que pensamos en la reproducción. Reproducirse no es solo tener hijos, puesto que desde el punto de vista evolutivo, lo importante no reside únicamente en cuántos descendientes directos produce un individuo, sino cuántas copias de sus genes logran atravesar el tiempo. Y esas copias no viajan sólo en los hijos. También viajan a través de sus nietos, hermanos, sobrinos, primos y otros parientes.
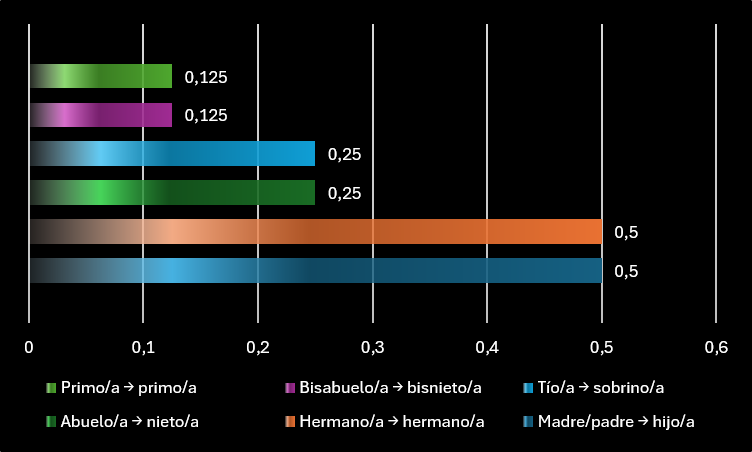
Un padre o una madre comparten, de media, el 50 % de su material genético con cada hijo. Es decir, cada hijo representa una transmisión directa de la mitad de su herencia genética. Pero esa transmisión no termina ahí. Si ese hijo sobrevive, crece y tiene a su vez descendencia, parte de esos genes continúan avanzando hacia la siguiente generación.
Un abuelo o una abuela comparten, de media, el 25 % de sus genes con cada nieto. A simple vista, puede parecer menos relevante que la relación entre madre e hijo, pero la evolución no opera solo en porcentajes aislados. Si una abuela ya no puede tener más hijos propios, pero su ayuda aumenta la supervivencia de varios nietos, reduce la carga de una hija, permite que esa hija tenga nuevos embarazos o mejora las probabilidades de que los niños lleguen a adultos, su influencia genética puede seguir siendo enorme. No transmite sus genes pariendo, sino ayudando a que sobrevivan quienes ya los portan.
Esta lógica se conoce como selección de parentesco. Su principio básico es sencillo: una conducta costosa puede verse favorecida por la evolución si el beneficio que produce en un pariente, multiplicado por el grado de parentesco con ese pariente, supera el coste que paga quien ayuda.
Dicho de otro modo, ayudar a un hijo “pesa” más que ayudar a un nieto, porque el parentesco genético es mayor. Pero ayudar a varios nietos, o mejorar de forma decisiva la supervivencia de una familia entera, puede compensar esa diferencia.
La evolución, por tanto, no se limita exclusivamente a contar nacimientos, también valora la supervivencia y los vínculos entre generaciones.
En términos cuantitativos, una ayuda evolutivamente costosa puede compensar si el beneficio para los parientes, ponderado por el parentesco, supera el coste de ayudar.
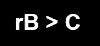
Donde r es el parentesco genético, B el beneficio para el pariente y C el coste para quien ayuda. Esta formulación se conoce como regla de Hamilton y es fundamental para entender por qué pueden evolucionar conductas de cooperación entre parientes. La abuela posreproductiva no estaría “fuera” de la reproducción por así decirlo, sino situada en otro estrato de la misma. Si bien no genera nuevos descendientes directos, sí que puede aumentar la probabilidad de que sobrevivan los descendientes ya existentes.
El precio de nacer: riesgos en el parto.
El nacimiento humano no constituye únicamente una prueba para la cría. También expone a la madre a hemorragias, infecciones, lesiones de los tejidos, y complicaciones derivadas de un parto prolongado u obstruido. La medicina contemporánea ha reducido enormemente estos peligros allí donde existen controles prenatales, personal especializado, transfusiones, antibióticos y capacidad para practicar una cesárea de urgencia. Sin embargo, el riesgo no ha desaparecido y continúa revelando hasta qué punto el parto supone un episodio crítico de la reproducción humana.
Por contextualizar, en pleno siglo XXI, y concretamente en 2023, murieron en el mundo unas 260.000 mujeres durante el embarazo, el parto o las semanas posteriores. La razón de mortalidad materna fue de 197 muertes por cada 100.000 nacidos vivos, aunque esta cifra oculta una desigualdad enorme: alrededor de 10 de cada 100.000 en los países más desarrollados, frente a 346 en aquellos con rentas más bajas. La mayoría de estas muertes podrían evitarse mediante una atención obstétrica adecuada y oportuna.
El riesgo afecta igualmente a la cría. En 2024 murieron aproximadamente 2,3 millones de recién nacidos durante sus primeros 28 días de vida, lo que corresponde a una mortalidad neonatal mundial de 17,2 por cada 1.000 nacidos vivos. La prematuridad, los episodios intraparto (entre ellos la asfixia y el traumatismo), las infecciones y la neumonía concentran la mayor parte de estas pérdidas. A ellas se suman los aproximadamente 1,9 millones de mortinatos registrados en 2023: fetos que murieron a partir de la semana 28 de gestación, más de cuatro de cada diez durante el propio trabajo de parto.
Estas cifras actuales no representan directamente el riesgo que afrontaron nuestras antepasadas. Ya que todo ello depende profundamente del acceso a la sanidad, la nutrición, las condiciones sociales y la calidad de la asistencia. Sin embargo, muestran que incluso en una especie dotada de tecnología médica, el intervalo que rodea al nacimiento sigue siendo uno de los momentos más vulnerables de la vida de la madre y del hijo.
El canal del parto
La dificultad del parto humano no depende únicamente del tamaño de la cabeza. El canal obstétrico tampoco forma un conducto recto y uniforme. Su entrada, su porción media y su salida presentan diámetros máximos orientados en direcciones diferentes. Por ello, la cabeza fetal no puede atravesarlo manteniendo una única posición: debe flexionarse y rotar mientras desciende.
En la presentación más habitual, el feto se encuentra longitudinalmente, con la cabeza dirigida hacia abajo y el mentón flexionado sobre el pecho. La cabeza suele introducirse en el estrecho superior con su eje mayor orientado de manera transversal u oblicua. A medida que desciende, gira para adaptar sucesivamente sus diámetros a las zonas más amplias del canal. La cabeza sale entonces por extensión: aparece primero la parte posterior del cráneo y, después, la frente, la cara y el mentón. Una vez liberada, vuelve a girar para alinearse con los hombros, que también deben rotar antes de atravesar la salida pélvica. La rotación fetal no es absolutamente exclusiva del ser humano, pero en nuestra especie es extraordinariamente frecuente y forma parte central del mecanismo normal del parto.
Esto es lo que se conoce como posición occipitoanterior, es decir, cuando la parte posterior de su cabeza queda orientada hacia el pubis materno, mientras que su cara mira hacia el sacro y la espalda de la madre. El recién nacido emerge así mirando en dirección contraria a ella, con su propia espalda orientada generalmente hacia la parte anterior del cuerpo materno.
Etapas del parto.
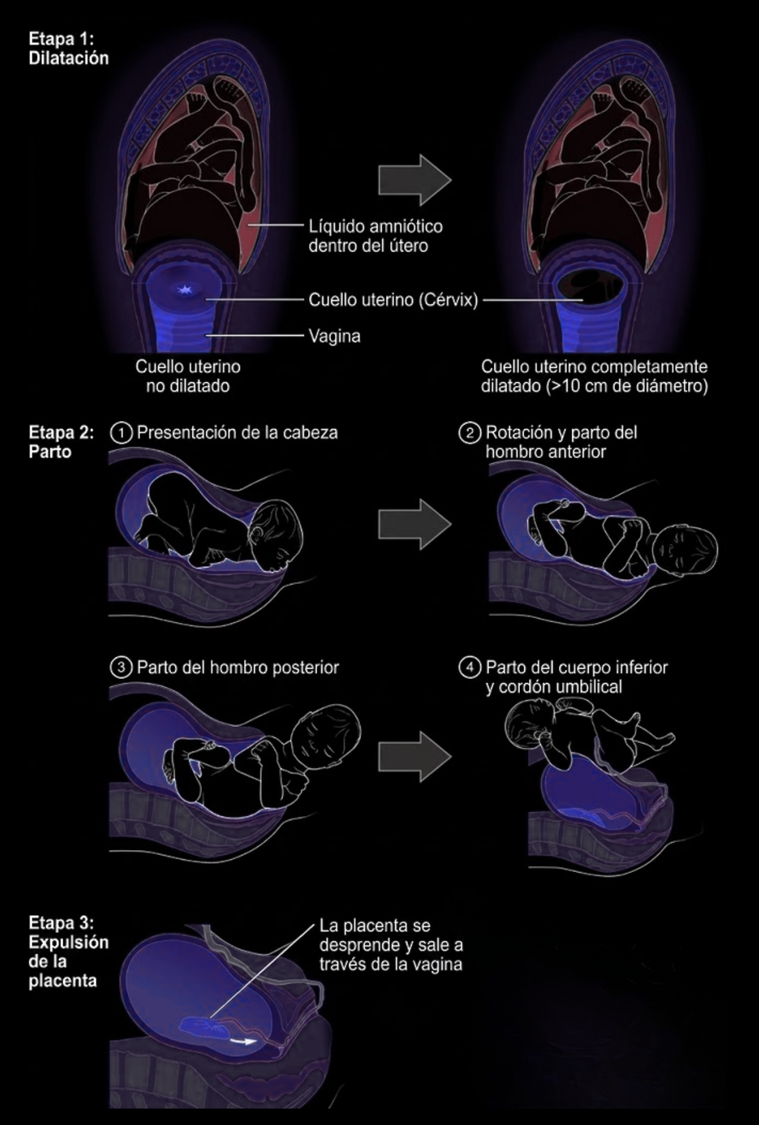
Esta disposición dificulta que la madre pueda controlar por sí misma la expulsión. Al inclinarse y extender los brazos hacia el periné, no puede ver con claridad la cara del recién nacido ni guiar fácilmente la salida de la cabeza y los hombros. Además, tirar de la cría desde esa posición podría forzar el cuello o desplazarla en sentido contrario a la curvatura natural que sigue el cuerpo al abandonar el canal. Una segunda persona, situada frente a la salida, puede sostener la cabeza, controlar la velocidad de expulsión, comprobar el estado del cordón, liberar los hombros y recibir al recién nacido antes de que caiga. De hecho, esta configuración anatómica ha sido propuesta como una de las bases evolutivas de la asistencia al parto humano.
Frente a otros mamíferos
En otras especies, como algunas entre los monos no humanos, la relación entre la cabeza neonatal y la pelvis materna permite un tránsito menos complejo. Con frecuencia, la cría emerge en posición occipitoposterior, es decir, mirando hacia la madre. Desde esa orientación, la hembra puede alcanzar al recién nacido, sujetarlo y dirigirlo hacia su cuerpo con mayor facilidad y seguridad. Sin embargo, este hecho no debe convertirse en una regla absoluta: algunos monos y grandes simios también muestran rotaciones fetales, presentaciones occipitoanteriores, partos difíciles e incluso intervención de otros individuos. Los chimpancés, por ejemplo, pueden presentar mecanismos de rotación semejantes en algunos aspectos a los humanos.
Tampoco conviene afirmar de forma categórica que todas las demás madres mamíferas paran necesariamente en soledad. El aislamiento es frecuente en numerosas especies, pero la asistencia social al nacimiento no es enteramente exclusiva de nuestra especie. Por ejemplo, en bonobos se ha observado que otras hembras permanecen junto a la parturienta, la protegen, inspeccionan la zona genital e incluso realizan gestos dirigidos a sujetar al recién nacido durante su salida. Estas madres podían completar el parto por sí mismas, lo que sugiere que la sociabilidad alrededor del nacimiento no depende únicamente de una imposibilidad anatómica de parir sin ayuda.
La diferencia humana reside más bien en la combinación de factores. El estrecho ajuste entre la cabeza y los hombros fetales y el canal materno, la necesidad habitual de rotación, la orientación con la que emerge el recién nacido, el riesgo de hemorragia y la vulnerabilidad inmediata de la cría convirtieron la presencia de otras personas en una ventaja considerable. A ello se añadieron la protección, el apoyo emocional, la transmisión de experiencia y la capacidad para actuar ante una complicación.
En prácticamente todas las sociedades humanas conocidas, el parto tiende por ello a ser un acontecimiento asistido. Existen excepciones culturales en las que parir sola se ha considerado una demostración de fortaleza, pero incluso en esos grupos la asistencia es habitual, especialmente durante los primeros nacimientos. La matrona, la madre, la hermana, la abuela u otras mujeres experimentadas no constituyen únicamente una presencia ceremonial: representan una respuesta social frente a un proceso anatómicamente exigente y potencialmente peligroso.
Esto sugiere que el parto humano pudo contribuir a reforzar una característica más amplia de nuestra estrategia reproductiva, ya que la reproducción dejó de ser una tarea estrictamente individual. La ayuda comenzaba antes incluso de que la cría hubiera abandonado por completo el cuerpo de su madre. Después vendrían la lactancia, el transporte, la alimentación, la protección y años de aprendizaje. El carácter social del nacimiento anticipaba, en cierto modo, el carácter cooperativo de toda la crianza humana.
El precio de nacer: los compromisos biomecánicos de la pelvis humana
Para complementar nuestra visión sobre la reproducción humana, conviene entender el precio que hemos pagado como especie. Un compromiso, algo perverso, que ha hecho la naturaleza entre las ventajas de nuestra anatomía, y el riesgo intrínseco al parto y la crianza prolongada. En resumidas cuentas, en nuestra especie, nacer es difícil. Algo que no suele ser así en la naturaleza, ni siquiera entre la inmensa mayoría de mamíferos, donde la madre asume una cantidad ingente de recursos tanto en la fecundación como en la crianza.
Además, las crías humanas son muy vulnerables hasta que llegan a la adolescencia. Pues nacen en un estado de profunda dependencia y requieren años de alimentación, protección y aprendizaje. Esta prolongada infancia se relaciona con nuestro lento desarrollo, con el intenso crecimiento cerebral posnatal y con la complejidad de las habilidades que deben adquirirse. El ajuste entre el feto y el canal del parto forma parte del problema, pero no constituye su única explicación.
Durante nuestra evolución convergieron, por tanto, diferentes presiones evolutivas. Por un lado, comenzamos a caminar erguidos, liberando las manos (esenciales para la manipulación fina), y por el otro, contamos con grandes cabezas para albergar nuestros cerebros. Esto último hizo que las crías presentaran cabezas grandes, hombros relativamente anchos, y un cerebro que requiere crecimiento.
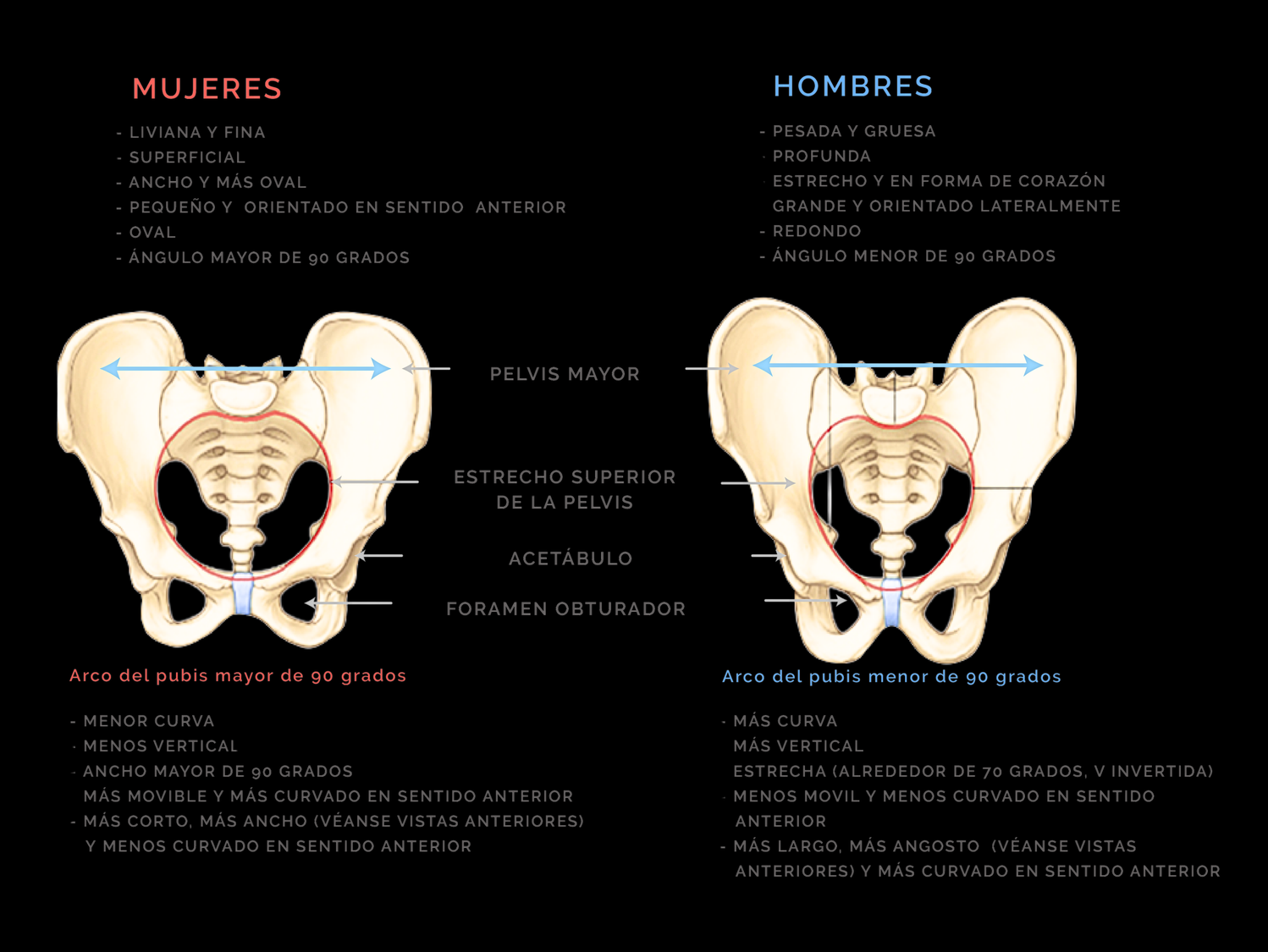
Para dar cabida a todas estas presiones evolutivas, la pelvis de la mujer se tuvo que reconfigurar. Un trámite biomecánico que ha de otorgar la integridad necesaria, propia para una estructura que soporta todo nuestro cuerpo y protege órganos internos, pero que además permita el parto de crías con cabezas grandes.
La pelvis femenina presenta diferencias anatómicas respecto a la masculina debido a su adaptación para el embarazo y el parto. Al ser más ancha y menos profunda, incrementa el espacio en la pelvis menor, facilitando así el paso de la cabeza del bebé al nacer.
La morfología obstétrica de la pelvis femenina puede imponer determinadas exigencias biomecánicas. Sin embargo, la magnitud y naturaleza de estos costes son más complejas de lo que propone el modelo clásico del dilema obstétrico. Una ampliación del canal del parto no implica necesariamente un aumento equivalente de la distancia entre las articulaciones de la cadera. La pelvis puede modificar su geometría interna y sus diámetros obstétricos sin alterar en la misma proporción la mecánica externa de la locomoción.
Según el modelo biomecánico estático clásico, una mayor distancia entre la cadera y la línea de acción de la fuerza de reacción del suelo aumentaría el momento externo que deben contrarrestar los abductores. Esto exigiría una mayor producción de fuerza muscular. Sin embargo, durante la locomoción real, el desplazamiento del centro de masas, la dirección de la fuerza de reacción y la cinemática de la extremidad modifican continuamente esos brazos de momento.
Como se podría deducir, una pelvis estrecha aumentaría la eficiencia locomotora, mientras que una pelvis ancha incrementaría la capacidad del canal de parto, reduciendo el riesgo de un parto obstruido. Mantener un canal de parto espacioso fue probablemente muy importante a partir del Pleistoceno Medio, cuando el tamaño del cerebro comenzó a aumentar rápidamente en el género Homo, afectando el tamaño craneal del neonato. Sin embargo, minimizar el costo locomotor también puede ser crucial para la aptitud reproductiva femenina.
No obstante, y al contrario de lo que se creía, estudios recientes han demostrado que las mujeres no necesariamente son menos eficientes al caminar o correr que los hombres.
Se sabe que las mujeres en sociedades cazadoras-recolectoras viajan regularmente 5 km o más al día, a menudo cargando un peso considerable, y además deben mantener recursos energéticos adecuados para el embarazo y la lactancia. Se cree que estas demandas evolutivas contrapuestas sobre la pelvis femenina son, al menos en parte, responsables de la dificultad del parto humano moderno y de la aparición de la desproporción cefalopélvica. Este escenario de compromiso, a menudo denominado el "dilema obstétrico", tiene consecuencias importantes para comprender el proceso del parto humano, la inversión materna y el desarrollo infantil.
Un desafío biomecánico único de la marcha bípeda humana es equilibrar el cuerpo sobre una sola extremidad de apoyo al caminar y correr. Debido a que la articulación de la cadera se encuentra a cierta distancia de la línea media del cuerpo, la pelvis tiende a rotar alejándose del lado de apoyo durante el soporte unilateral. Los músculos abductores de la cadera (glúteo medio, glúteo menor y tensor de la fascia lata) contrarrestan esta rotación produciendo una fuerza opuesta en la pelvis, redirigiendo así el centro de masa del cuerpo para mantener el equilibrio mediolateral. Para mantener el equilibrio en la articulación de la cadera, el momento externo que actúa sobre ella en el plano mediolateral debe ser contrarrestado por un momento interno igual y opuesto, generado principalmente por los músculos abductores de la cadera.
Bajo el modelo biomecánico estático estándar, la fuerza del músculo abductor se determina por la magnitud de la fuerza externa (la fuerza de reacción del suelo GRF) y la ventaja mecánica efectiva (EMA) de los abductores de la cadera: la relación entre el brazo de palanca del músculo abductor de la cadera (r) y el brazo de palanca de la GRF (R). Dado que este modelo asume que la GRF pasa casi verticalmente a través del centro de masa del cuerpo en la mitad del apoyo de la marcha, se piensa que R es aproximadamente igual a la mitad de la anchura biacetabular. La anchura biacetabular se mide tradicionalmente desde el aspecto más interno del acetábulo. Sin embargo, dado que la rotación de la articulación de la cadera ocurre alrededor del centro de la cabeza femoral, la anchura biacetabular se define como la distancia entre los centros de las cabezas femorales, una medida más relevante desde el punto de vista biomecánico para comprender la función de los abductores de la cadera. Si la mitad de la anchura biacetabular es un buen indicador de R durante la locomoción, como asume el modelo estándar, entonces aumentar este diámetro pélvico disminuirá la EMA de los abductores y, asumiendo que no hay cambios en la GRF ni en r, requerirá más fuerza muscular y una mayor energía metabólica para mantener la estabilidad pélvica durante la fase de apoyo unilateral al caminar y correr. Por lo tanto, el dilema obstétrico predice que la mayor anchura pélvica en las mujeres, asociada con las demandas de dar a luz a bebés de cerebro grande, compromete la EMA de los abductores de la cadera y resulta en una locomoción menos eficiente en las mujeres en comparación con los hombres.
El estudio de Warrener et al. (2015) cuestionó uno de los pilares biomecánicos clásicos del dilema obstétrico: la idea de que una pelvis femenina más ancha facilita el parto, pero empeora necesariamente la eficiencia de la marcha y la carrera. En la muestra estudiada, una mayor anchura pélvica no se asoció con un mayor coste energético de locomoción. Por tanto, el canal del parto humano no parece estar limitado simplemente porque una pelvis más ancha haría a las mujeres menos eficientes al desplazarse.
Comparación entre el modelo estático clásico y el cálculo dinámico mediante dinámica inversa. El primero aproxima el brazo externo a partir de la anchura biacetabular; el segundo lo calcula empleando la cinemática de los segmentos corporales y las fuerzas registradas durante la locomoción.
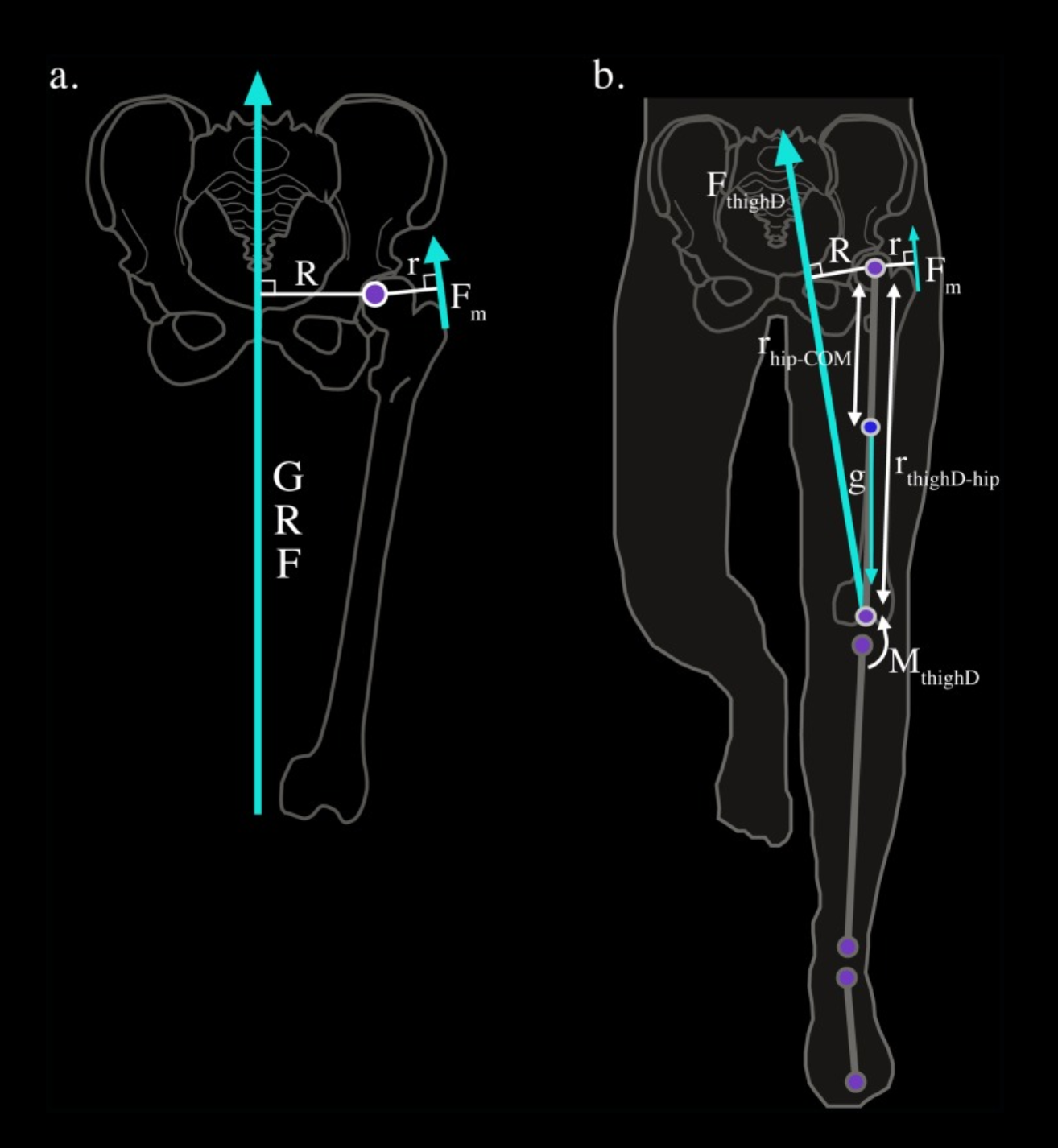
La dinámica inversa ofrece un enfoque alternativo para estimar la producción dinámica de fuerza de los abductores de la cadera al caminar y correr. Este método integra los datos de la fuerza de reacción del suelo (GRF) de una plataforma de fuerza que informa sobre las aceleraciones del centro de masa de todo el cuerpo con los datos cinemáticos de cada segmento de la extremidad inferior. Brevemente, el método modela la extremidad inferior como una serie de segmentos vinculados en la que el pie, la espinilla y el muslo actúan como cuerpos rígidos que interactúan en articulaciones sin fricción. Dado que se conocen tanto la fuerza externa que actúa sobre el modelo de la extremidad (la GRF) como la cinemática de cada segmento, los momentos alrededor de cada articulación responsables de las aceleraciones angulares observadas se pueden resolver mediante ecuaciones de Newton-Euler.
Dimensiones antropométricas y variables biomecánicas de la muestra estudiada. Crédito: https://pmc.ncbi.nlm.nih.gov/articles/PMC4356512/
Glosario de términos
- Brazo de momento r del abductor: Distancia perpendicular (brazo de palanca o de momento) medida desde el centro de rotación de la articulación de la cadera hasta la línea de acción de los músculos abductores (como el glúteo medio).
- Cuello femoral: Zona ósea estrecha y cilíndrica del fémur que conecta la cabeza femoral con la diáfisis.
- Anchura biespinosa: La distancia lineal entre las espinas isquiáticas de la pelvis. Es una métrica muy relevante en antropología y obstetricia porque delimita uno de los diámetros más estrechos del canal de parto.
- Estrecho inferior medio-lateral de la pelvis: Medida en el plano medio-lateral de la apertura inferior de la pelvis (la salida del canal del parto).
- Anchura biacetabular: distancia entre los centros de rotación de ambas articulaciones de la cadera, determinados a partir de los centros de las cabezas femorales. En este estudio se emplea como una medida biomecánicamente más relevante que la distancia entre los bordes internos de los acetábulos.
- Diámetro de la cabeza femoral: El diámetro de la esfera superior del fémur. Su dimensión suele correlacionarse estrechamente con el tamaño y el peso que soporta el esqueleto.
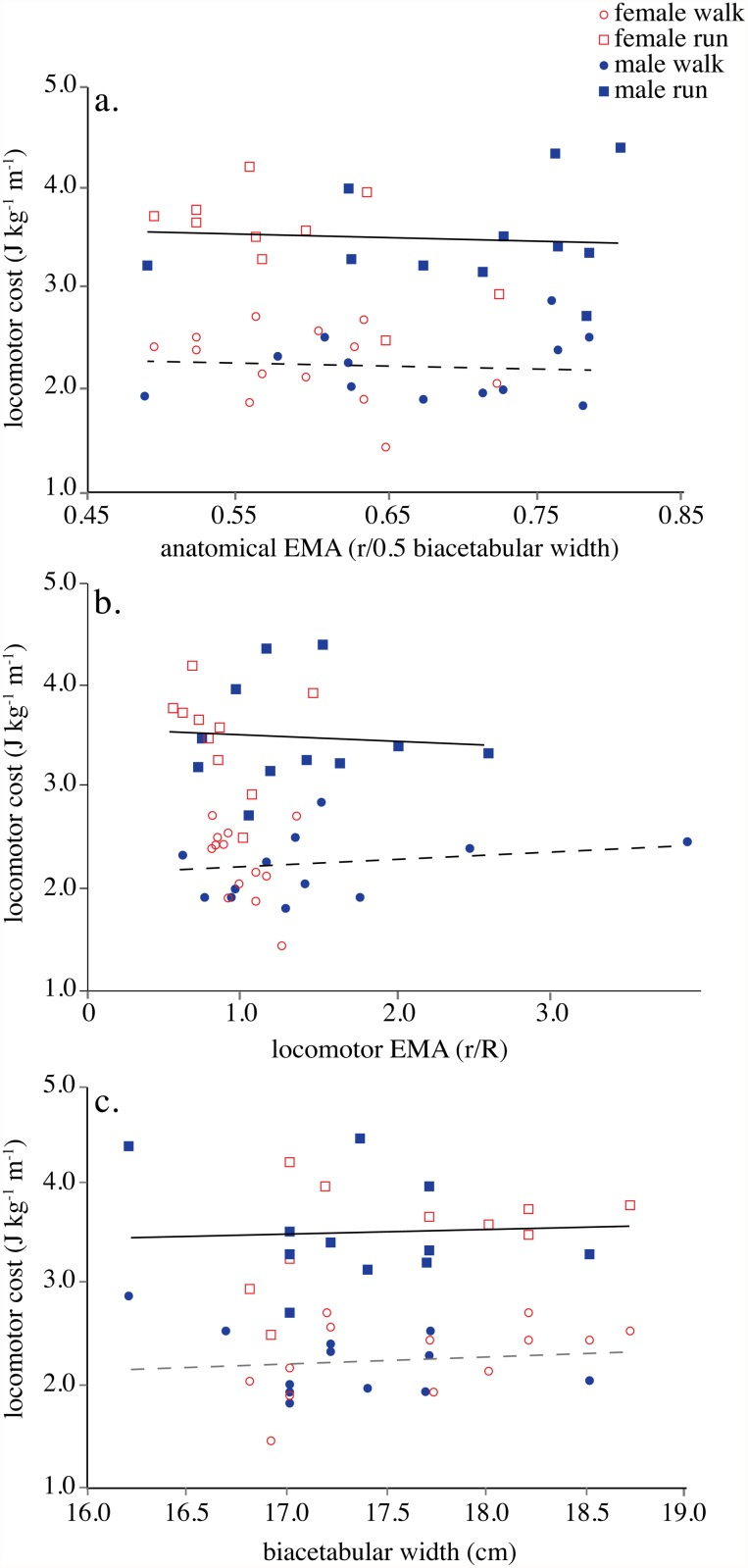
- VME/EMA anatómica: estimación estática de la ventaja mecánica de los abductores, calculada como la relación entre su brazo de momento, r, y la mitad de la anchura biacetabular, utilizada como aproximación del brazo externo R.
La ventaja mecánica efectiva (EMA) de los abductores de la cadera medida anatómica y dinámicamente frente al costo locomotor.
- Círculo rojo: mujer andando
- Cuadrado rojo: mujer corriendo
- Círculo azul: hombre andando
- Cuadrado azul: hombre corriendo
Una mayor demanda local de los abductores no implicó una mayor demanda locomotora del organismo en su conjunto.
Costo metabólico de la activación de los abductores de la cadera y costo locomotor neto en hombres y mujeres.
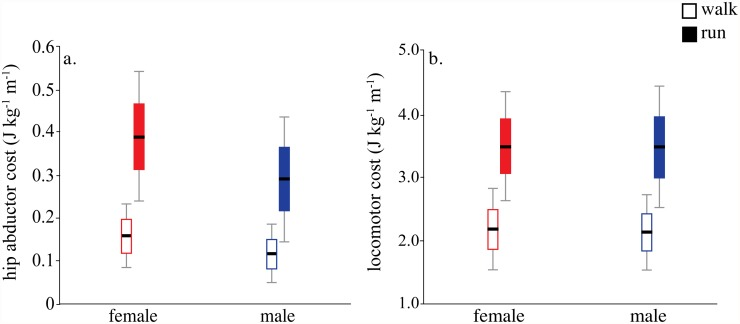
En el contexto de este estudio, las mujeres de la muestra presentaron, en promedio, una menor ventaja mecánica anatómica de los abductores. Como consecuencia, el coste estimado de activación de esta musculatura fue aproximadamente un 35 % mayor. Sin embargo, los abductores representaban una fracción relativamente pequeña del gasto locomotor total, por lo que esta diferencia local no se tradujo en una menor economía global al caminar o correr.
Eso conlleva que sus abductores principalmente glúteo medio, glúteo menor y tensor de la fascia lata deben generar más fuerza para producir un mismo momento estabilizador sobre la pelvis. Las mujeres presentaron brazos de momento abductores más cortos y una menor ventaja mecánica anatómica. Como consecuencia, el coste estimado de activación de los abductores fue aproximadamente un 35 % mayor. Sin embargo, estos músculos representaban una proporción reducida del gasto locomotor total, por lo que la economía global de la marcha y la carrera no difirió significativamente entre hombres y mujeres.
Ahora bien, lo que se concluye es que esto no se debía a una mayor distancia entre las articulaciones de la cadera, sino principalmente a que las mujeres tienen brazos de momento abductores más cortos. La longitud del brazo de momento se relacionaba especialmente con:
- la longitud del cuello femoral,
- la anchura biilíaca,
- el tamaño corporal general.
La longitud del cuello femoral explicó aproximadamente un 62 % de la variación del brazo de momento abductor, y la anchura biilíaca alrededor del 51 %.
El estudio, aunque revelador, presenta varias limitaciones. La muestra biomecánica era relativamente pequeña y estaba formada por corredores recreativos jóvenes y físicamente activos. El coste metabólico se midió sobre cinta a velocidades prefijadas, mientras que la cinemática y las fuerzas se registraron sobre una pista interior con plataforma de fuerza. Por tanto, quedaron fuera situaciones como la locomoción prolongada, el terreno irregular, el transporte de cargas, la fatiga, el embarazo y parte de la actividad de la musculatura del tronco.
En cualquier caso, esto nos invita a pensar una cosa, y es que, aunque este estudio pueda tener ciertas limitaciones, sí que debilita en cierto modo la clásica premisa que afirma que la pelvis femenina no puede ser más ancha porque una pelvis ancha encarecería energéticamente la locomoción bípeda. Estos resultados no lo respaldan, pero sí que deben existir otras restricciones que expliquen por qué la selección natural no produjo canales del parto todavía más amplios. Para explicar esto se han propuesto otras posibilidades, como la estabilidad del suelo pélvico, otros aspectos del rendimiento locomotor, la termorregulación, entre otras.
Si la eficiencia locomotora global no explica por sí sola las dimensiones relativamente limitadas del canal del parto, el suelo pélvico ofrece otro posible compromiso. Esta estructura, formada por músculos y tejidos conectivos, debe sostener las vísceras, el útero gestante y las presiones generadas en la cavidad abdominal, pero también deformarse intensamente durante el nacimiento.
Mediante modelos de elementos finitos, Stansfield y colaboradores observaron que, al aumentar el tamaño radial del suelo pélvico mientras se mantenían constantes su grosor y la presión aplicada, aumentaban su descenso, el estiramiento de los tejidos y las tensiones internas. Un canal pélvico más amplio obliga, en este sentido, a suspender los tejidos sobre una abertura mayor, lo que puede comprometer su capacidad de soporte y contribuir al riesgo de disfunciones como la incontinencia o el prolapso.
Aumentar el grosor del suelo pélvico mejora su resistencia a la deformación, pero no elimina completamente el efecto de una abertura mayor. Además, un tejido más grueso y rígido exige una presión intraabdominal superior para distenderse durante el parto. Se configura así un segundo compromiso: un suelo pélvico suficientemente resistente para sostener los órganos, pero también suficientemente deformable para permitir el nacimiento. La selección natural no habría actuado sobre una única oposición entre caminar y parir, sino sobre un sistema de exigencias anatómicas parcialmente incompatibles.
El efecto de la variación en el radio estandarizado del suelo pélvico, R, sobre (A) el desplazamiento absoluto del suelo pélvico, (B) el esfuerzo de Von Mises y (C) Estiramiento. R se expresa como un múltiplo de la dimensión del modelo base. Un valor de R de 1 equivale al tamaño promedio de la población femenina, y aproximadamente el 95% de esta población se encuentra dentro del rango de R de 0.84 a 1.16 (±2 DE). El grosor del suelo pélvico y la presión se mantuvieron constantes (6 mm y 4 kPa, respectivamente). El desplazamiento, el esfuerzo y el estiramiento se midieron por separado para los compartimentos anterior y posterior (símbolos "x" azul y "+" verde, respectivamente), así como para la solución analítica aplicada a una membrana circular (línea discontinua roja). Para dimensiones intermedias del suelo pélvico (0.8 < R < 1.1), el compartimento anterior mostró un aumento aproximadamente lineal en el desplazamiento con una tasa similar a la de la solución analítica, mientras que exhibió un aumento sustancial no lineal en el desplazamiento para suelos pélvicos más grandes que el promedio. El desplazamiento en el compartimento posterior aumentó a una tasa ligeramente más lenta que la solución analítica hasta R = 0.95. A partir de ahí, el aumento en el desplazamiento posterior fue similar al de la solución analítica y lo superó para tamaños muy grandes (R > 1.4).
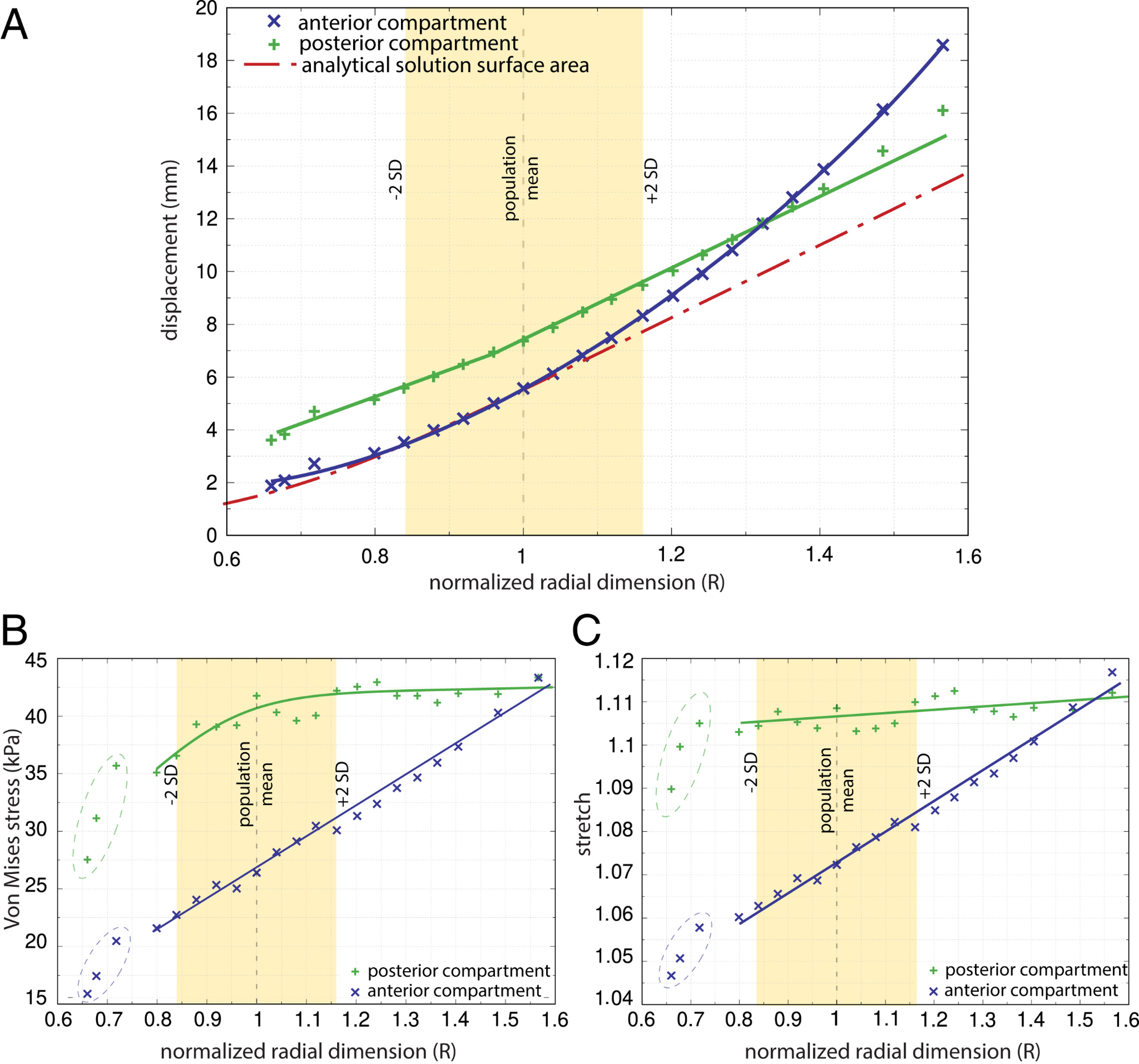
Tamaño y grosor del suelo pélvico como compromiso biomecánico. Los modelos de elementos finitos muestran que, manteniendo constante el grosor del tejido y la presión aplicada, el aumento del radio del suelo pélvico produce un mayor desplazamiento hacia abajo. Por el contrario, incrementar su grosor aumenta la rigidez y reduce la deformación. Sin embargo, un suelo pélvico más grueso también requiere una mayor presión intraabdominal para deformarse durante el parto. La evolución habría tenido que equilibrar, por tanto, la amplitud del canal obstétrico con la capacidad de sostener las vísceras y permitir el nacimiento.
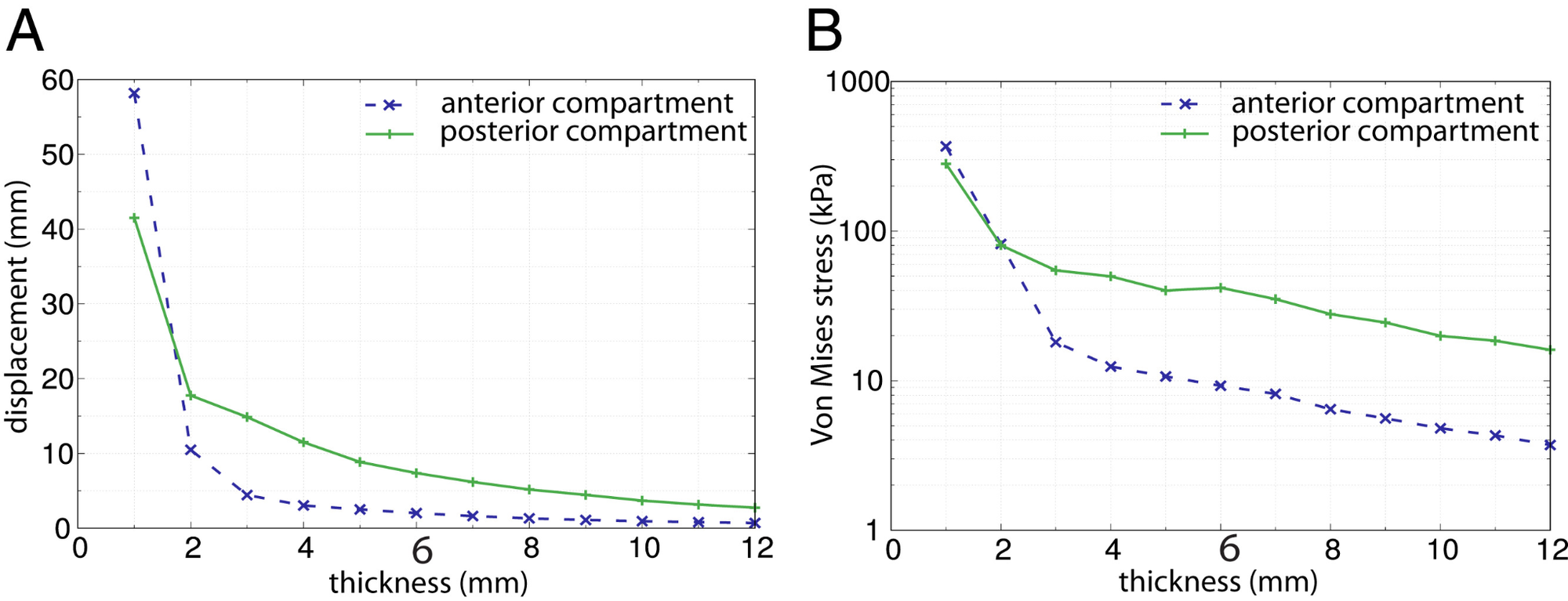
Nota: resultados obtenidos mediante modelización por elementos finitos; no representan mediciones clínicas directas de continencia o prolapso.
Por otro lado, un estudio de modelización musculoesquelética publicado en 2026 matizó estos resultados. Al modificar virtualmente la anchura pélvica en un ±20 %, la demanda energética de los abductores cambió aproximadamente un ±10 %, mientras que el efecto sobre el coste locomotor corporal total se mantuvo en torno al ±1 %. Otros grupos musculares compensaban buena parte del incremento local. Los autores plantean, por tanto, que el posible coste evolutivo de una pelvis más ancha podría haber estado relacionado menos con el gasto energético global que con la fatiga localizada o la capacidad de mantener esfuerzos prolongados, aunque esta posibilidad no fue medida directamente.
La evolución no pudo resolver estas presiones eliminando el riesgo, pero lo distribuyó de modo que satisfaciera, en mayor o menor medida, todos los compromisos en juego. La pelvis femenina amplió determinadas dimensiones del canal obstétrico sin sufrir una penalización locomotora global apreciable, pero conservó un ajuste estrecho entre madre y feto y mantuvo importantes exigencias sobre la musculatura estabilizadora y el suelo pélvico.
Ahora bien, tras el parto, además, comenzaba una segunda dificultad: alimentar, proteger y enseñar durante años a una cría profundamente dependiente. En ese contexto, la reproducción humana difícilmente podía descansar exclusivamente sobre la madre. La ayuda del padre, de las abuelas y del resto del grupo pudo transformar una limitación en una estrategia de crianza cooperativa.
Fertilidad y edad
Una ventana que se estrecha
La fertilidad femenina no se mantiene constante hasta la menopausia para desaparecer de forma repentina. El envejecimiento reproductivo comienza mucho antes y afecta tanto a la cantidad como a la calidad de los ovocitos. A diferencia de lo que ocurre con la producción continua de espermatozoides, la reserva ovárica femenina se establece antes del nacimiento y disminuye de manera irreversible: de unos 6–7 millones de ovocitos durante la vida fetal se pasa a aproximadamente 1–2 millones al nacer, entre 300.000 y 500.000 en la pubertad, cerca de 25.000 alrededor de los 37 años y alrededor de un millar al aproximarse la menopausia. La mayor parte no llega a ovular, sino que desaparece mediante procesos naturales de degeneración folicular.
Definición de ovocitos, atresia y degeneración folicular
- Ovocitos: células germinales femeninas inmaduras contenidas en los folículos de los ovarios. Durante su maduración completan distintas fases de la meiosis y, tras la ovulación y la fecundación, pueden dar lugar a un embrión. En sentido estricto, la célula liberada durante la ovulación humana es un ovocito secundario, no un óvulo completamente maduro.
- Degeneración folicular o atresia folicular: proceso natural por el cual los folículos ováricos que no llegan a completar su maduración se deterioran y son reabsorbidos por el organismo. La gran mayoría de los folículos desaparece mediante atresia; solo una pequeña proporción alcanza la ovulación. Este proceso contribuye a la disminución progresiva de la reserva ovárica con la edad.
- Atresia: proceso natural de degeneración y reabsorción de una estructura o tejido que deja de desarrollarse.
Esta reducción cuantitativa se acompaña de un deterioro progresivo de la calidad ovocitaria. Con la edad aumentan los errores durante la división cromosómica del ovocito, de modo que se forman con mayor frecuencia embriones con un número anómalo de cromosomas. Muchos de estos embriones no llegan a implantarse, otros originan pérdidas gestacionales tempranas y una pequeña proporción puede continuar su desarrollo con alguna alteración cromosómica. Por ello, la edad de la mujer constituye uno de los principales determinantes de la probabilidad de concepción, aborto espontáneo y nacimiento vivo.
La fertilidad natural alcanza sus valores máximos durante la veintena y los primeros años de la treintena. Después comienza un descenso gradual que se hace más acusado a partir de mediados de los treinta y, especialmente, alrededor de los 37 años. A los 40 años, la fertilidad relativa es aproximadamente la mitad de la observada entre el final de la veintena y el comienzo de la treintena. Esto no significa que el embarazo natural sea imposible a partir de esa edad, sino que normalmente requiere más tiempo y que una proporción creciente de concepciones no culmina en un nacimiento.
El descenso de la fertilidad no solo se manifiesta en una menor probabilidad de quedar embarazada. También aumenta la posibilidad de perder la gestación. Entre los 20 y los 30 años, aproximadamente entre el 9 y el 17 % de los embarazos clínicamente reconocidos terminan en una pérdida temprana. La proporción se sitúa alrededor del 20 % a los 35 años, del 40 % a los 40 y puede alcanzar aproximadamente el 80 % a los 45. Cerca de la mitad de las pérdidas gestacionales tempranas se relacionan con anomalías cromosómicas embrionarias. Estas cifras son estimaciones poblacionales y no permiten predecir el resultado de un embarazo individual.
Además, una vez establecida la gestación, la edad materna avanzada se asocia con una mayor frecuencia de diabetes gestacional, hipertensión, preeclampsia (una de las formas en que la hipertensión arterial puede presentarse en el embarazo), alteraciones placentarias, parto prematuro, restricción del crecimiento fetal, cesárea y muerte fetal. El incremento no aparece súbitamente al cumplir 35 años, puesto que se trata de un fenómeno continuo que se vuelve más evidente entre los 35 y los 39 años y más pronunciado a partir de los 40. La frontera de los 35 años es principalmente una referencia clínica e histórica para intensificar el asesoramiento y la vigilancia, no una división entre embarazos «seguros» y «peligrosos».
La muerte fetal tardía sigue siendo poco frecuente en términos absolutos, pero su probabilidad aumenta con la edad y con la prolongación del embarazo. Según el consenso, al alcanzar las 41 semanas el riesgo relativo de muerte fetal es aproximadamente tres veces mayor entre las gestantes de 40 años o más que entre las menores de 35. Este aumento contribuye a que en algunos casos se recomiende una vigilancia más estrecha y se valore finalizar la gestación antes de que se prolongue excesivamente.
También aumenta el riesgo para la madre, aunque la magnitud absoluta depende profundamente de la salud previa, la obesidad, la hipertensión, la diabetes, la atención prenatal y la capacidad del sistema sanitario para responder a una emergencia. Como ejemplo, en Estados Unidos la mortalidad materna registrada en 2024 fue de 16,5 muertes por cada 100.000 nacidos vivos entre las mujeres de 25 a 39 años y de 62,3 por cada 100.000 entre las de 40 años o más. Esta medida incluye las muertes durante el embarazo, el parto y el postparto, por lo que no representa exclusivamente el riesgo de morir durante la expulsión del bebé.
La relación entre edad y seguridad reproductiva no es completamente lineal. Aunque las mujeres jóvenes poseen, en promedio, una reserva ovárica abundante y ovocitos de buena calidad, los embarazos durante la adolescencia también presentan riesgos superiores a los observados al comienzo de la edad adulta. Las madres de entre 10 y 19 años muestran una mayor incidencia de eclampsia, endometritis e infecciones sistémicas que las de 20 a 24 años; sus hijos presentan con mayor frecuencia prematuridad, bajo peso al nacer y estados neonatales graves. En este caso intervienen tanto factores biológicos como nutricionales, sociales y asistenciales.
Por tanto, el periodo de menor riesgo medio se sitúa aproximadamente durante la veintena y los primeros años de la treintena. Sin embargo, hablar de una «edad perfecta» resulta engañoso, puesto que la fertilidad y el riesgo no cambian mediante escalones rígidos, sino a través de una transición gradual, diferente en cada mujer. La edad de 35 años no constituye un precipicio reproductivo, del mismo modo que conservar ciclos menstruales regulares no garantiza que la cantidad y calidad de los ovocitos permanezcan intactas.
Desde la perspectiva evolutiva, esta progresiva reducción de la fertilidad resulta especialmente relevante. Con la edad, la reproducción deja de ofrecer el mismo balance entre beneficio y riesgo. Disminuye la probabilidad de concebir, aumenta la frecuencia de pérdidas gestacionales y se elevan los peligros para la madre y para la cría. En una especie de desarrollo lento, donde los hijos siguen dependiendo durante años de alimento, protección y aprendizaje, la muerte de una mujer durante un nuevo embarazo podría comprometer no solo al recién nacido, sino también a los descendientes que todavía necesitaban de ella.
Llegado cierto punto, invertir en una nueva gestación podría resultar menos ventajoso que contribuir a la supervivencia y reproducción de los hijos ya nacidos.
Aquí comienza el papel evolutivo de la abuela:
Una mujer que ha dejado atrás su etapa reproductiva directa, pero que todavía puede aumentar la transmisión de sus genes ayudando a sus descendientes y nietos.

Como ya se ha mencionado, desde una perspectiva evolutiva, la reproducción no se limita a engendrar hijos propios. Un individuo también puede favorecer la transmisión de sus genes ayudando a sobrevivir y reproducirse a sus parientes. Una abuela comparte, en promedio, una cuarta parte de sus genes con cada nieto; por tanto, mejorar su alimentación, protección o supervivencia puede producir un beneficio evolutivo indirecto.
Esta es la idea central de la hipótesis de la abuela. La prolongación de la vida femenina más allá de la fertilidad habría podido ser favorecida porque las mujeres mayores aportaban recursos, conocimiento y cuidados al grupo familiar. Su ayuda podía aumentar la supervivencia de los nietos, aliviar la carga materna y permitir que sus hijas tuvieran nuevos hijos en intervalos más cortos.
La abuela no sustituía a la madre, sino que ampliaba su capacidad reproductiva. Allí donde una mujer sola encontraba límites para alimentar, transportar y proteger a varias crías dependientes, la presencia de una cuidadora experimentada podía transformar el equilibrio entre el coste de la crianza y el número de descendientes que llegaban a sobrevivir.
Esto no significa que la menopausia surgiera únicamente porque los embarazos tardíos fueran peligrosos. El riesgo obstétrico ayuda a explicar por qué la reproducción a edades avanzadas puede perder rentabilidad, pero la hipótesis de la abuela requiere además que la supervivencia posreproductiva genere beneficios suficientes para los descendientes. La selección no habría favorecido simplemente “dejar de tener hijos”, sino seguir viviendo y siendo útil después de dejar de tenerlos.
El precio de nacer no terminaba en el parto. Continuaba durante años de dependencia. Precisamente por eso, en nuestra especie, una mujer podía contribuir tanto a su legado evolutivo ayudando a criar como volviendo a parir.
La hipótesis de la madre
La hipótesis de la madre propone que la menopausia pudo ser favorecida porque, a partir de cierta edad, una mujer obtenía más éxito evolutivo protegiendo e invirtiendo en los hijos que ya tenía que intentando tener nuevos hijos.
En los seres humanos, los hijos permanecen dependientes durante muchos años. Si una madre de edad avanzada vuelve a quedar embarazada, afronta simultáneamente:
- mayor riesgo de morir durante el embarazo o el parto;
- mayor riesgo de aborto, muerte fetal o alteraciones congénitas;
- un nuevo periodo de lactancia y crianza intensiva;
- competencia energética entre el recién nacido y los hijos que todavía dependen de ella.
Por tanto, una nueva reproducción no sólo pone en riesgo a la madre y al nuevo hijo, sino que también amenaza indirectamente la supervivencia de todos los hijos dependientes que ya existen. Los modelos de esta hipótesis incorporan precisamente que la muerte materna durante la crianza reduce sustancialmente la supervivencia infantil.
Los modelos teóricos muestran que la hipótesis puede producir una menopausia adaptativa cuando coinciden dos condiciones:
- un aumento acusado con la edad de la mortalidad materna asociada a la reproducción;
- un aumento importante de la muerte fetal, las alteraciones congénitas y el fracaso reproductivo.
En esas circunstancias, abandonar la reproducción y mantener una vida posreproductiva puede proporcionar una ventaja selectiva.
Sin embargo, los resultados empíricos no son concluyentes. Un análisis de poblaciones humanas premodernas de Finlandia y Canadá no encontró pruebas suficientes de que el riesgo de muerte materna y la protección de los hijos existentes, por sí solos, hubieran generado una selección suficientemente intensa para explicar la menopausia humana.
La principal limitación es que la hipótesis de la madre puede explicar razonablemente la interrupción de la reproducción, pero tiene más dificultades para explicar una fase posreproductiva de varias décadas. Una vez que los hijos propios alcanzan la independencia, el beneficio estrictamente maternal disminuye. Para explicar la longevidad posterior deben añadirse otros mecanismos, como:
- la ayuda a los nietos;
- la transferencia de recursos a hijos adultos;
- el conocimiento ecológico acumulado;
- la reducción del conflicto reproductivo entre generaciones;
- la cooperación de padres, abuelos y hermanos mayores.
Conviene aclarar que la hipótesis de la abuela y la hipótesis de la madre explican fenómenos diferentes. La hipótesis de la madre explica principalmente por qué podría ser ventajoso dejar de reproducirse antes de morir, mientras que la hipótesis de la abuela intenta explicar además por qué sería ventajoso vivir durante muchos años después de la menopausia.
Evidencia de la hipótesis de la abuela. El papel de la abuela en números

Con tal de aportar evidencia, se han seleccionado algunos artículos cuyas conclusiones apoyan en mayor o menor medida la hipótesis de la abuela:
Límites a los beneficios biológicos de una vida posreproductiva prolongada en las mujeres.
El trabajo usa registros parroquiales históricos de la Finlandia preindustrial, entre 1731 y 1895, con datos de nacimientos, muertes, matrimonios, hijos, ocupaciones y linajes familiares. La muestra principal incluye 5.815 nietos, 1.034 abuelas maternas y 1.003 abuelas paternas, con el estado de ambas abuelas conocido. Algunos de los datos barajados en el estudio se encuentran en la tabla siguiente:
En lo que respecta a la población, generalmente el hijo mayor solía heredar la granja, la dispersión era baja y, aunque tanto abuelas maternas como paternas podían vivir cerca, las abuelas paternas eran las que más típicamente podían convivir con los nietos.
En cuanto a los resultados obtenidos, la disponibilidad de nietos a los que una mujer podía ayudar aumentaba desde los 40 años, alcanzaba un máximo en los primeros 60, y después descendía rápidamente. Hacia los 75 años, la mayoría de los nietos ya habían nacido, por lo que disminuía la oportunidad de aumentar la fitness ayudando a nuevos nietos.
Además, la mortalidad femenina aumentaba mucho justo cuando esas oportunidades empezaban a caer:
El resultado más favorable aparece en las abuelas maternas de 50–75 años. Su presencia se asocia con una mayor supervivencia (sus odds de sobrevivir eran un 29,5 % mayores) de los nietos entre los 2 y 5 años, es decir, después del destete, una etapa especialmente vulnerable. Ahora bien, esto no significa que la supervivencia aumentara directamente en 29,5 puntos porcentuales. Las odds son una medida comparativa distinta del porcentaje de supervivencia. Por ejemplo, si en el grupo de referencia sobreviviera el 95 % de los niños, un efecto de esta magnitud elevaría la supervivencia estimada aproximadamente hasta el 96 %, no hasta el 124,5 %.
Esto es una evidencia directa a favor de la hipótesis de la abuela, ya que una mujer post-reproductiva puede fomentar su linaje no teniendo más hijos, sino ayudando a que sobrevivan sus nietos. Como contraparte se observa que, para los nietos de 2 a 5 años, la presencia de abuelas paternas no se asocia significativamente con mayor supervivencia.
Otro detalle que vislumbra el estudio es que las abuelas paternas de +75 pueden ser perjudiciales, asociándose con una menor supervivencia de los nietos entre el nacimiento y los 2 años. En cambio, las abuelas maternas de más de 75 años no tuvieron un efecto negativo estadísticamente significativo. La interpretación propuesta por los autores es que las abuelas paternas muy mayores, al ser más propensas a convivir en el hogar, podrían convertirse en una carga de recursos, cuidados o estrés familiar, especialmente en un contexto preindustrial de alta mortalidad infantil y recursos limitados.
Otro detalle, que no es baladí, es que la salud de la abuela importa tanto como la edad. Como no tenían historiales médicos, los autores usaron el tiempo restante hasta la muerte de la abuela como proxy de salud, y se encontraron que una abuela paterna que moriría en el plazo de un año se asociaba con menor supervivencia infantil, mientras que una abuela materna que estaba a 3 o más años de morir seguía teniendo un efecto positivo sobre los nietos de 2-5 años.
El estudio sugiere una especie de ventana óptima de valor adaptativo de la abuela. Una abuela post-reproductiva relativamente sana, especialmente materna y de 50–75 años, puede aumentar la supervivencia de los nietos. Pero cuando la edad avanza demasiado, disminuyen tanto las oportunidades de ayudar como la capacidad fisiológica o material de hacerlo.
Se podría entender, tras los datos arrojados que la selección pudo favorecer una vida larga tras la menopausia mientras existieran beneficios netos para los descendientes, pero estos beneficios se debilitan cuando la abuela pasa a competir por cuidados o recursos.
El uso de la distancia geográfica como un indicador potencial de ayuda en la evaluación de la hipótesis de la abuela
Este estudio analiza si la presencia de una abuela materna viva mejora el éxito reproductivo de sus hijas y, especialmente, si ese beneficio disminuye cuando madre e hija viven más alejadas.
Los autores emplearon registros genealógicos y parroquiales de los colonos franceses del valle del río San Lorenzo, en Quebec, durante los siglos XVII y XVIII. La muestra principal comprendió 740 abuelas maternas y 2.274 hijas, con distancias entre ellas de hasta aproximadamente 325 km. Un hecho importante es que se realizaron comparaciones dentro de una misma familia, por ejemplo entre hermanas cuya madre estaba viva o había muerto cuando ellas desarrollaron su vida reproductiva. De este modo se reducía, aunque no se elimina completamente, la influencia de factores genéticos, familiares y ambientales compartidos.
La distancia geográfica se utilizó como un indicador indirecto de la posibilidad de ayuda: se supone que una abuela cercana puede proporcionar con mayor facilidad cuidados infantiles, alimentos, trabajo doméstico, apoyo durante embarazos y partos o asistencia en situaciones de enfermedad. El estudio no registró directamente esas ayudas.
La presencia de la abuela materna se asoció con un incremento medio de aproximadamente:
- 2,1 hijos nacidos adicionales por hija.
- 1,1 hijos adicionales que sobrevivieron hasta los 15 años.
Por tanto, las mujeres con una madre viva no solo tuvieron más descendientes, sino también un mayor número de hijos que alcanzaron una edad utilizada por los autores como aproximación a la supervivencia hasta la madurez.
Este último resultado debe interpretarse cuidadosamente: el estudio demuestra un aumento del número absoluto de hijos supervivientes, pero no implica necesariamente que la abuela elevara en la misma proporción la probabilidad individual de supervivencia de cada niño. Parte del efecto puede deberse a que las hijas tuvieron más nacimientos.
Entre las mujeres cuyas madres estaban vivas, el beneficio disminuía al aumentar la separación geográfica. A mayor distancia entre madre e hija:
- Descendía el número total de hijos nacidos.
- Descendía el número de hijos que sobrevivían hasta los 15 años, es decir, el éxito reproductivo vital.
- Aumentaba la edad de la hija al iniciar la reproducción.
Distancias geográficas y rasgos de la historia de vida.
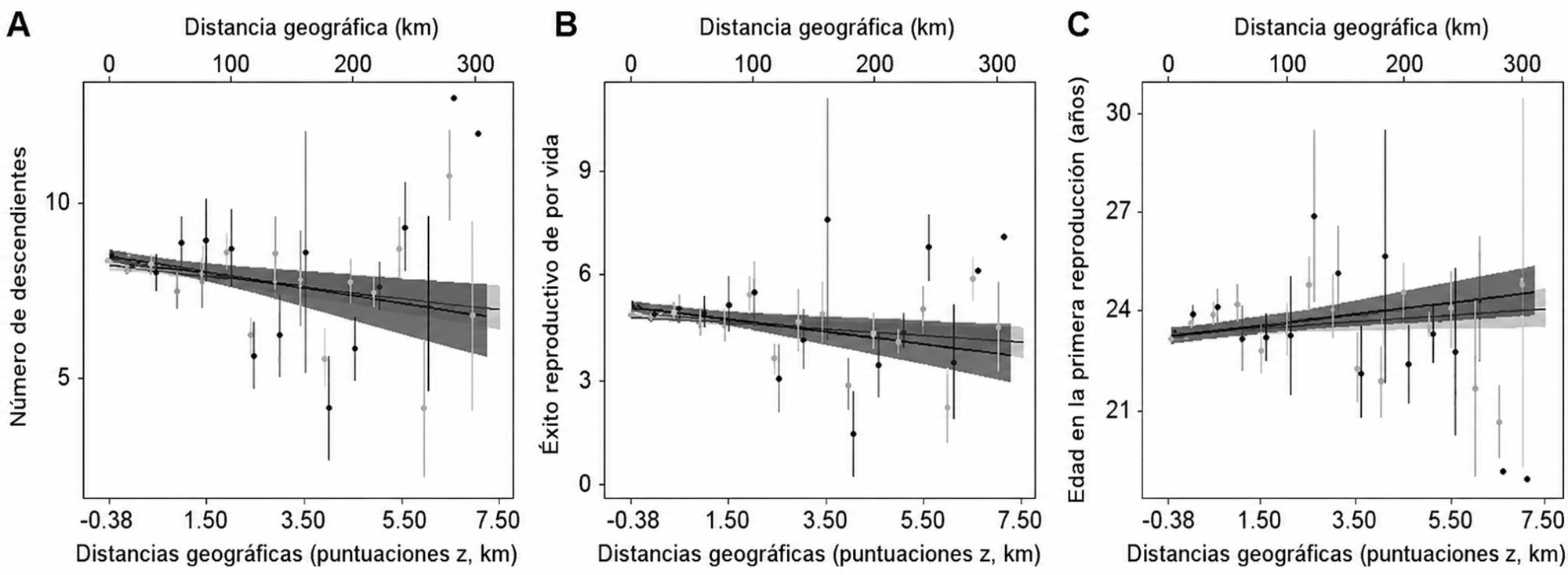
En consecuencia, una abuela viva pero distante se asociaba con un beneficio mucho menor que una abuela viva y cercana.
Esto constituye el resultado más relevante del artículo: no es solamente la longevidad de la abuela lo que importa, sino la posibilidad efectiva de interactuar y colaborar con sus descendientes.
Los resultados son compatibles con la hipótesis de la abuela, y el patrón espacial encontrado refuerza esta interpretación, ya que si el efecto dependiera únicamente de una constitución genética familiar favorable o de que las mujeres longevas pertenecieran a familias más sanas, la distancia entre madre e hija no debería reducirlo de manera tan clara. Que el beneficio disminuya con la separación es coherente con un mecanismo basado, al menos parcialmente, en la ayuda presencial. No obstante, la distancia es únicamente un indicador indirecto. Los registros históricos no permiten saber qué hacía realmente cada abuela, cuánto tiempo dedicaba a sus nietos ni qué recursos proporcionaba.
Además, vivir cerca puede reflejar otros factores como mayor cohesión familiar, similitud socioeconómica, acceso compartido a tierras o recursos o menor migración de las hijas.
Evolución de las historias de vida: El papel de las abuelas en el espacio y el tiempo.
Este artículo no presenta un experimento nuevo, sino que interpreta conjuntamente los dos estudios históricos vistos previamente: el realizado con poblaciones de la antigua Nueva Francia y el otro con registros parroquiales finlandeses.
El límite espacial
La presencia de una abuela no produce automáticamente una ventaja. Para que exista un beneficio evolutivo, la abuela debe tener acceso real a sus descendientes y capacidad para prestar ayuda.
Cuando aumenta la separación geográfica:
- disminuye la posibilidad de cuidar directamente a los niños;
- disminuye la transferencia cotidiana de alimentos, trabajo o conocimientos;
- disminuye la capacidad de liberar a la hija de parte de la carga reproductiva;
- y, en consecuencia, se reduce el beneficio reproductivo atribuible a la abuela.
La distancia se interpreta, por tanto, como una aproximación a la oportunidad efectiva de ayudar. El artículo rechaza implícitamente una formulación demasiado simple de la hipótesis: no basta con que la abuela esté viva, sino que debe poder intervenir materialmente en la vida de sus descendientes.
El límite temporal
La eficacia de la ayuda también cambia con la edad. El artículo señala que el beneficio asociado a la presencia de la abuela comienza a disminuir aproximadamente hacia los 70 años.
Los autores relacionan este descenso con dos procesos simultáneos:
- Disminuyen los receptores potenciales de ayuda. A medida que la mujer envejece, sus nietos suelen ser mayores y menos dependientes, por lo que existen menos oportunidades de mejorar su supervivencia mediante cuidados directos.
- Disminuye la capacidad de ayudar. El deterioro físico, la enfermedad y el aumento de la mortalidad reducen la cantidad y eficacia de la ayuda que puede proporcionar la abuela.
Por tanto, la vejez posreproductiva no mantiene indefinidamente el mismo valor evolutivo. Existe una etapa en la que la mujer todavía puede aportar más recursos de los que consume, seguida de otra en la que su contribución puede disminuir e incluso convertirse en una demanda de cuidados para la familia.
Esquema del cronograma de los costes (valores negativos de W) y beneficios (valores positivos de W) de la aptitud inclusiva para una mujer focal (F0) a lo largo de dos generaciones, basado de forma general en los hallazgos de la base de datos finlandesa. Las barras representan las historias de vida superpuestas de las mujeres F0 y F1; los periodos reproductivos y posreproductivos se muestran en verde y azul; los periodos de conflicto intergeneracional potencial sobre la reproducción y el rol de abuela se muestran en amarillo y morado, respectivamente.
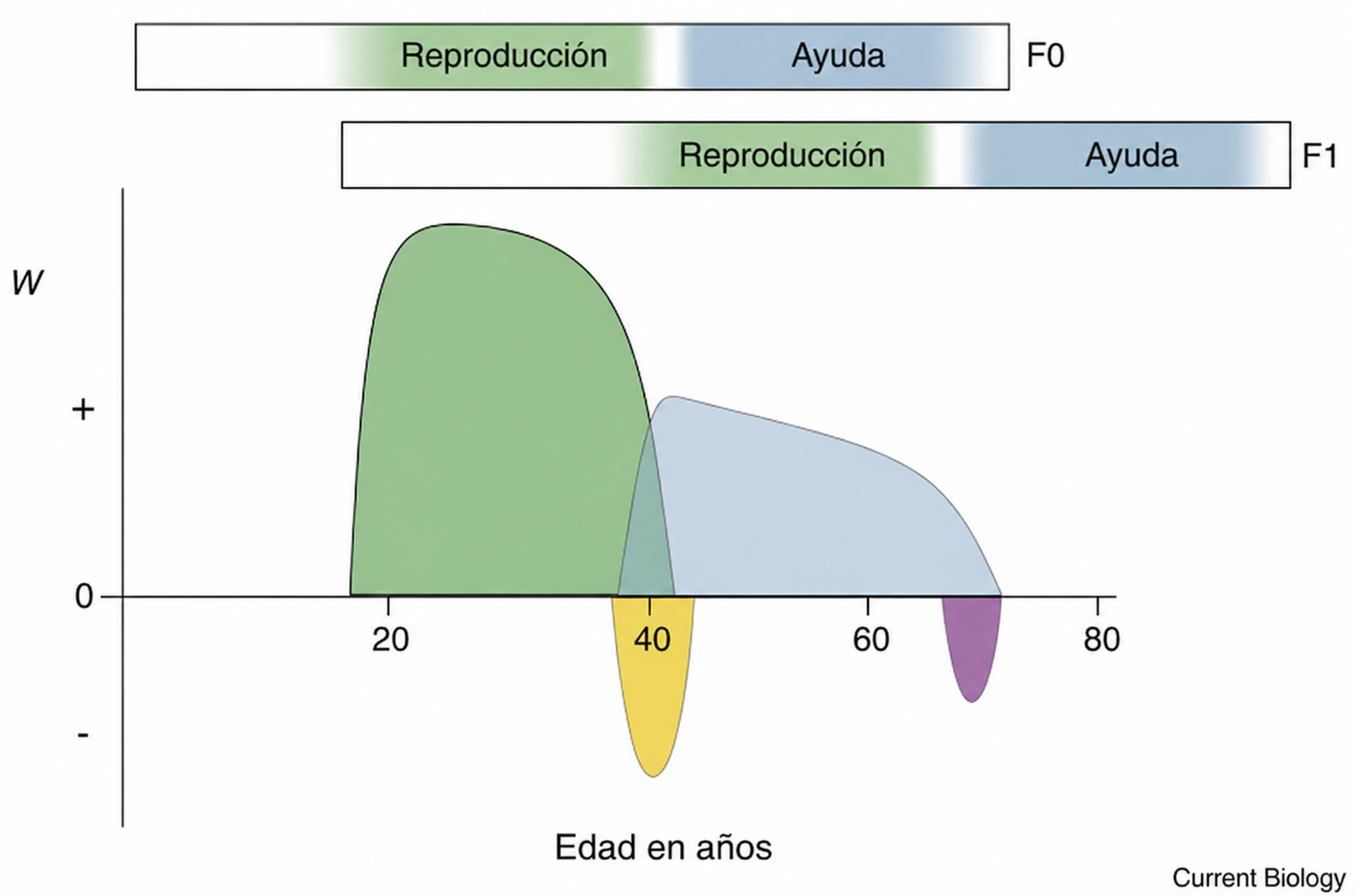
En esta tabla el eje horizontal representa la edad en años, desde el comienzo de la edad adulta hasta aproximadamente los 80 años. Por otro lado, el eje vertical representa la contribución a la aptitud inclusiva, indicada como W, donde por encima de cero aparecen los beneficios y por debajo de cero aparecen los costes.
La aptitud inclusiva incluye tanto:
- la aptitud directa, obtenida teniendo hijos propios;
- como la aptitud indirecta, obtenida ayudando a que sobrevivan y se reproduzcan otros familiares que comparten parte de sus genes.
Por otro lado, las dos barras superiores muestran las historias vitales superpuestas de dos generaciones:
- F0: la mujer focal, que posteriormente se convierte en abuela.
- F1: su hija.
En cada barra:
- el tramo verde representa la etapa de reproducción propia;
- el tramo azul representa la etapa de ayuda posreproductiva.
La hija F1 comienza su etapa reproductiva cuando F0 está terminando la suya. Después, F0 pasa de producir nuevos hijos a ayudar a F1 con sus descendientes.
Área verde: La gran superficie verde representa la aptitud obtenida por F0 al tener y criar a sus propios hijos. El beneficio aumenta al comenzar la vida reproductiva, alcanza un máximo durante las edades de mayor fecundidad y disminuye conforme se aproxima el final de la reproducción.
Esto no ha de interpretarse como una curva exacta de fecundidad, sino como una representación esquemática de que la reproducción propia es la principal fuente de aptitud durante la primera parte de la vida adulta.
Área azul: La superficie azul comienza aproximadamente cuando disminuye la reproducción propia. Representa los beneficios indirectos derivados de:
- alimentar o cuidar a los nietos,
- ayudar a la hija durante la reproducción,
- permitirle reducir el intervalo entre nacimientos,
- transmitir experiencia o conocimientos,
- y mejorar la supervivencia de los descendientes.
El área azul disminuye progresivamente porque, al avanzar la edad de F0, se reduce tanto su capacidad de ayudar como el número de nietos que todavía necesitan ayuda.
Área amarilla: La depresión amarilla situada alrededor de los 40 años representa el coste del conflicto reproductivo intergeneracional.
Este conflicto aparece cuando F0 continúa reproduciéndose al mismo tiempo que F1 inicia su propia reproducción. Ambas generaciones pueden competir por:
- alimentos;
- cuidados;
- espacio doméstico;
- trabajo de otros familiares;
- y otros recursos necesarios para criar a sus respectivos hijos.
La gráfica sugiere que dejar de reproducirse reduce ese solapamiento. La transición de F0 desde la reproducción hacia la ayuda evita parte de la competencia y transforma una relación potencialmente conflictiva en una relación cooperativa.
Área verde: La depresión morada, alrededor de los 70 años, representa lo que los autores denominan como «coste de la abuelidad tardía». No significa necesariamente que exista una disputa consciente entre la abuela y sus descendientes. Representa el momento en que:
- disminuye su capacidad de producir o prestar ayuda,
- aumentan su fragilidad y sus necesidades,
- requiere recursos o cuidados de generaciones más jóvenes,
- y queda un número menor de nietos dependientes a los que pueda beneficiar.
La misma mujer que anteriormente era una proveedora neta puede convertirse progresivamente en receptora neta de ayuda. Desde el punto de vista de la aptitud inclusiva, esto introduce un coste tardío que limita la ventaja de prolongar indefinidamente la vida.
De todo esto, se podría deducir que las abuelas pueden aumentar su aptitud inclusiva cuando tienen descendientes dependientes, viven suficientemente cerca de ellos y conservan la capacidad de proporcionar una ayuda efectiva. Por lo tanto, el efecto de la abuela depende de la ecología familiar y no constituye una ventaja universal e invariable.
La menopausia habría evitado una crisis energética en la mediana edad gracias al apoyo de hijos mayores aún dependientes y de los progenitores: un estudio de simulación.
En este artículo, el autor construye dos simulaciones deterministas. Para ello representa una población estacionaria dividida por edad y sexo. Para cada grupo calcula:
balance energético = producción alimentaria - gasto energético total
El consumo se estima mediante el gasto energético diario total, ajustado por edad, sexo, peso y talla. La producción depende de:
- fuerza física, aproximada mediante la masa corporal,
- habilidad de subsistencia adquirida con la edad,
- productividad máxima adulta,
- importancia relativa de fuerza y habilidad.
Este modelo permite observar si la producción conjunta de adultos, jóvenes y niños puede sostener al conjunto de la población. No representa familias concretas ni decide quién alimenta a quién: presupone que los excedentes producidos por unas personas pueden compensar los déficits de otras.
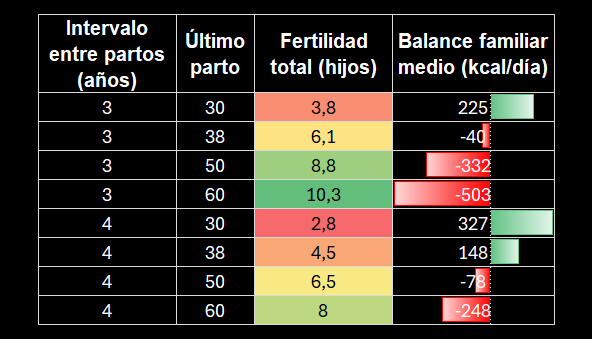
Para el modelo familiar, se supone una unidad formada por madre, padre e hijos dependientes, donde sus principales parámetros son:
Los hijos nacen regularmente cada tres o cuatro años y dejan de pertenecer a la unidad dependiente a los 20. Se supone que todas las mujeres comienzan a reproducirse a los 20 años y mantienen una fecundidad constante hasta una edad de último parto prefijada. Es importante entender que la edad del último parto no equivale exactamente a la menopausia fisiológica. El modelo modifica el momento en que cesan los nacimientos, pero no representa el agotamiento folicular, la disminución progresiva de la fecundidad, los abortos espontáneos ni el cese definitivo de la menstruación.
La particularidad humana es que las madres pueden tener un nuevo hijo aproximadamente cada tres años, mientras que cada descendiente tarda cerca de dos décadas en alcanzar la autosuficiencia completa.
Por tanto, una mujer puede tener simultáneamente:
- un lactante;
- uno o varios niños pequeños;
- niños mayores;
- adolescentes que todavía consumen recursos, aunque ya producen parte de ellos.
Al principio de la vida reproductiva, la producción de la pareja aumenta lentamente, pero el consumo crece con rapidez a medida que nacen hijos. Esto conduce a un déficit inicial. Durante la segunda década reproductiva, los hijos mayores comienzan a producir alimentos y reducen la carga neta, pero continúan naciendo niños si la reproducción no se detiene.
Para ello, se modela la productividad como:

donde:
- TEEprop representa la producción energética máxima como proporción del gasto energético diario total de un adulto.
- Se asignan valores de 0,4–1,6 para las mujeres y de 1–2,2 para los hombres.
- α expresa el peso relativo de la fuerza frente a la habilidad:
- 0,25: producción basada principalmente en la habilidad,
- 0,50: igual importancia de fuerza y habilidad,
- 0,750: producción basada principalmente en la fuerza.
La fuerza se aproxima mediante el peso corporal correspondiente a cada edad y sexo, expresado respecto al peso máximo de un varón adulto. A este valor se le aplica una función que provoca una caída rápida al aproximarse la edad máxima:

El parámetro b0, que determina la velocidad del declive durante la vejez, se fija en −0,15 para ambos sexos. La función pretende representar procesos como la pérdida de masa muscular y de densidad ósea.
La habilidad para obtener alimentos se representa mediante una función de aprendizaje, combinada con otra función que reduce la habilidad al aproximarse la edad máxima:
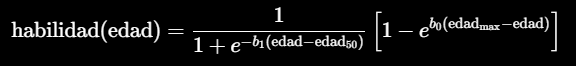
Aquí:
- b1 controla la rapidez con la que se adquieren las habilidades;
- edad50 es la edad a la que se alcanza el 50 % de la habilidad máxima.
El autor plantea tres trayectorias de aprendizaje:
Estas trayectorias se aplican por separado a hombres y mujeres. La productividad se considera nula en los niños menores de tres años.
En esencia, el modelo supone que la capacidad de producir alimentos depende simultáneamente de:
tamaño y fuerza corporal + conocimientos y destrezas acumulados − deterioro por la vejez
De los datos arrojados se puede deducir que la edad del último parto produce un intercambio muy claro entre número de hijos y sostenibilidad energética.
Resultados medios del modelo familiar. Crédito (adaptado): https://arxiv.org/pdf/2601.13442
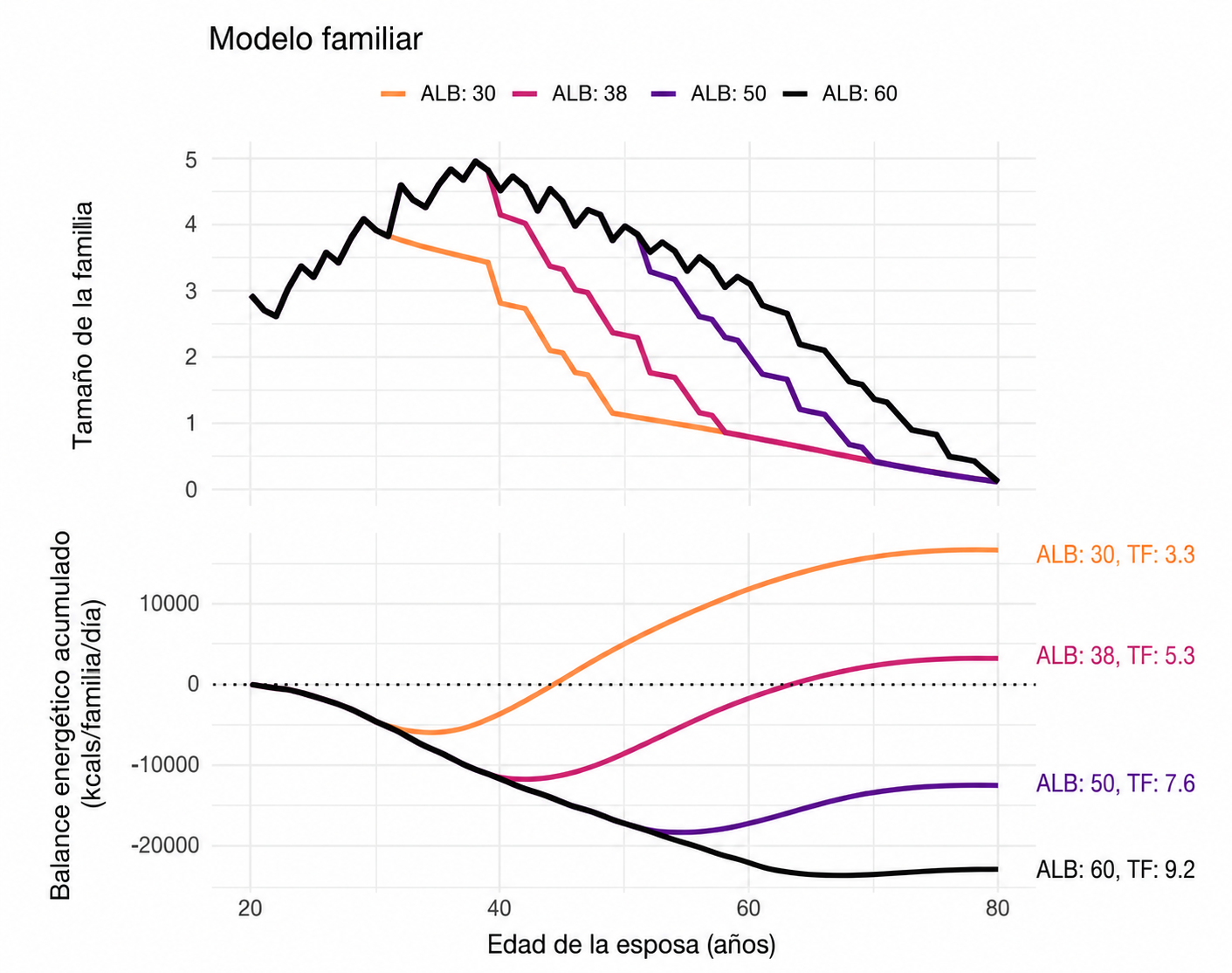
Modelo familiar. Superior: Tamaño medio de la familia según la edad de la esposa para diferentes valores de ALB (promediado entre los diferentes valores de IBI). Inferior: Suma acumulada media del balance energético familiar diario desde la AFB de la esposa hasta la esperanza de vida máxima, para diferentes valores de ALB (promediado en todo el espacio de parámetros). Los valores a los 80 años cercanos o superiores a cero indican que, en promedio, la familia acabó produciendo la suficiente energía excedente total para compensar sus déficits energéticos totales mediante transferencias de energía, mientras que los valores negativos indican que el excedente total fue inferior al déficit total. Nótese la compensación entre la fertilidad total (promediada para IBI = 3 e IBI = 4) y el balance energético para diferentes valores de ALB.
Notas sobre acrónimos
- ALB (Age of Last Birth): Edad al último nacimiento.
- AFB (Age of First Birth): Edad al primer nacimiento.
- IBI (Inter-Birth Interval): Intervalo entre nacimientos.
- TF (Total Fertility): Fertilidad total (o tasa de fecundidad total).
El consumo, la producción y el balance diario de energía de la esposa, el esposo y sus hijos dependientes a lo largo de la vida de la esposa, promediados en todo el espacio de parámetros. Balance = producción menos consumo.
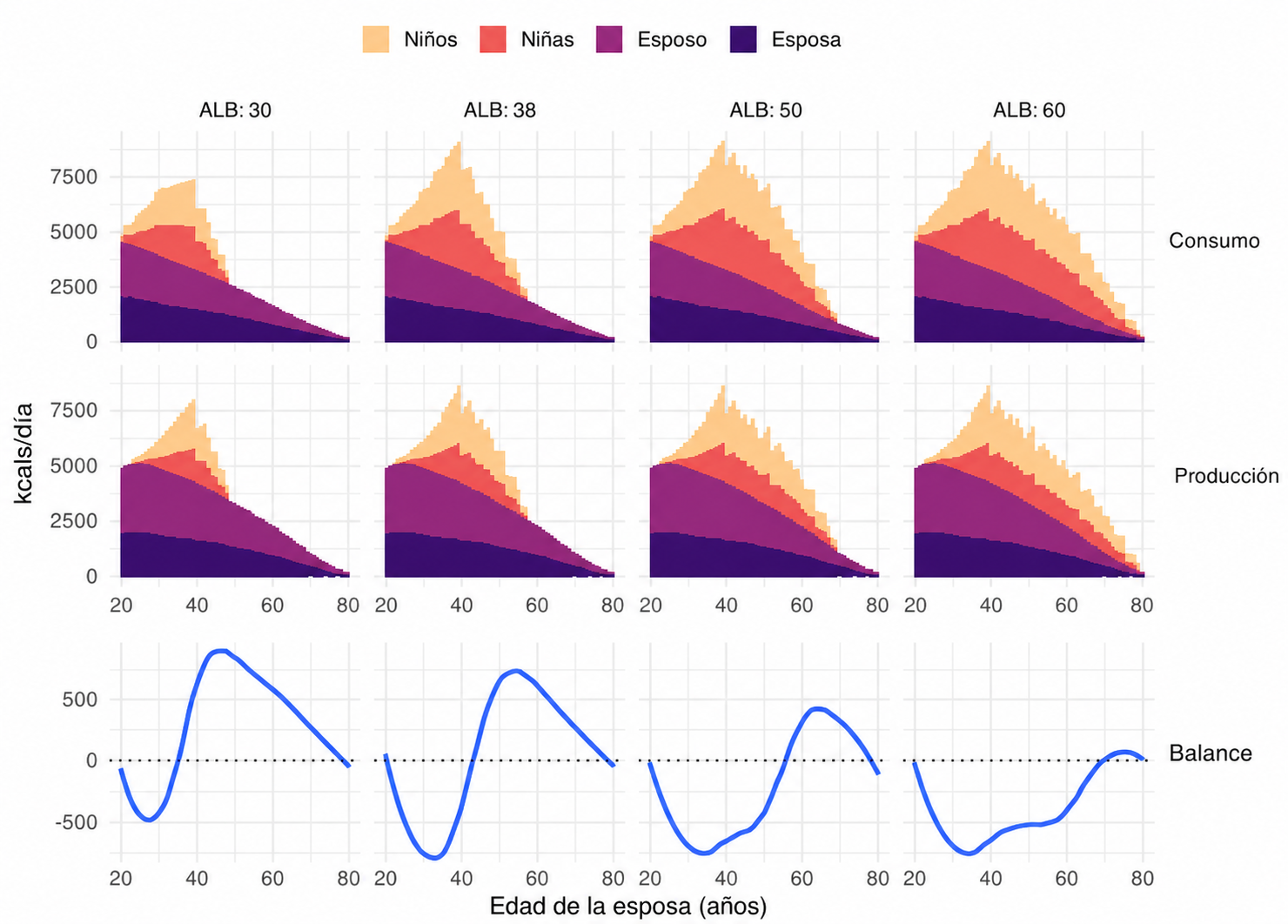
Los valores están promediados sobre todo el espacio de parámetros, por lo que no representan una sociedad ancestral concreta. Aun así, muestran una transición consistente: prolongar la reproducción hasta los 50 o 60 años aumenta la descendencia potencial, pero deja a las familias con déficits energéticos mucho mayores y más prolongados.
La edad de último parto cercana a los 38 años se encuentra en una posición especialmente interesante, ya que no maximiza ni la producción energética ni el número de hijos, pero constituye una especie de frontera de compromiso, ya que permite una fertilidad elevada, limita el tiempo durante el cual permanecen numerosos dependientes y deja una fase posterior de la vida en la que los padres pueden generar excedentes.
No puede afirmarse que sea el «óptimo evolutivo», porque el modelo no calcula éxito reproductivo inclusivo. Pero sí aparece como un compromiso energéticamente plausible entre cantidad y capacidad de inversión.
Por lo tanto, los resultados muestran que el ciclo vital humano genera una crisis energética previsible durante la mediana edad. La combinación de intervalos entre partos relativamente cortos y una dependencia juvenil cercana a dos décadas hace que las parejas acumulen varios descendientes que todavía consumen más de lo que producen. Cuando la reproducción se prolonga hasta los 50 o 60 años, las familias alcanzan una fertilidad mayor, pero permanecen más tiempo en déficit y disponen de menos años para generar excedentes. Por el contrario, detener los nacimientos alrededor de los 38–40 años limita la carga familiar y permite que el balance mejore conforme maduran los hijos.
Este resultado apoya una versión energética de la hipótesis de la madre: el cese reproductivo no sólo evitaría los riesgos crecientes de un nuevo embarazo, sino que protegería la inversión ya realizada en los descendientes existentes. La ventaja no estaría simplemente en tener menos hijos, sino en evitar que continúen incorporándose niños pequeños cuando los padres envejecen y todavía mantienen a varios dependientes. En este sentido, la menopausia favorecería la calidad y supervivencia potencial de la descendencia ya nacida frente a la producción indefinida de nuevos hijos.
El estudio también respalda tanto la hipótesis de la abuela como el modelo de capital incorporado de ambos sexos. Una vez finalizada la reproducción y reducida la carga de hijos dependientes, las mujeres y los hombres mayores pueden producir o conservar excedentes transferibles a hijos adultos y nietos. Los resultados constatan la necesidad de transferencias intergeneracionales, pero no permiten determinar si estas procedían principalmente de las abuelas, los abuelos, los padres, otros parientes o un sistema comunitario de reparto. Por ello, el artículo apoya mejor una hipótesis general de las generaciones ascendentes que una explicación exclusivamente centrada en la abuela.
A estas transferencias se suma un elemento que normalmente se pasa por alto, la producción juvenil. Los dependientes generaron, en promedio, alrededor del 60 % de sus propias necesidades energéticas, y su contribución resultó esencial para mantener el equilibrio poblacional en la mayor parte de los escenarios. Los niños mayores y adolescentes no serían, por tanto, receptores pasivos: primero reducen su propio déficit y posteriormente pueden ayudar a hermanos menores. Los adultos jóvenes sin descendencia también pueden producir excedentes, lo que resulta compatible con formas de crianza cooperativa, ayuda entre hermanos y prácticas como el servicio del novio a la familia de la esposa.
La hipótesis del padre ausente recibe apoyo parcial y debe reformularse. Prolongar la reproducción aumenta el número de hijos que quedarían dependientes si muriera el padre, pero la muerte materna también era frecuente. En las simulaciones, alrededor de los 50 años era casi tan probable que sobreviviera únicamente el marido como únicamente la esposa. La presión selectiva relevante habría sido, por tanto, evitar que la muerte de cualquiera de los progenitores dejara a varios niños pequeños sin suficiente apoyo, más que prevenir específicamente la ausencia del padre.
El trabajo plantea que la menopausia no habría evolucionado por un único beneficio aislado, sino como parte de un sistema integrado formado por cese reproductivo, inversión biparental, productividad creciente de los hijos, cooperación entre hermanos y transferencias de padres y abuelos. Este sistema permitiría compatibilizar una fertilidad relativamente elevada con una infancia prolongada y costosa. La menopausia actuaría así como un mecanismo que cierra la fase reproductiva antes de que la acumulación de dependientes impida sostener a la familia y ayudar a la generación siguiente.
No obstante, estas simulaciones identifican una presión energética plausible, no demuestran por sí solas el origen evolutivo de la menopausia. El modelo no evalúa directamente el envejecimiento de los ovocitos, los riesgos obstétricos, la competencia reproductiva entre parientes ni las explicaciones no adaptativas. Tampoco puede decidir entre la hipótesis de la abuela, el modelo biparental o el reparto comunitario, porque no especifica quién transfiere los recursos a quién. Sus conclusiones deben entenderse como evidencia de compatibilidad funcional, no como una prueba definitiva de causalidad evolutiva.
Cuidar también es reproducirse
La hipótesis de la abuela ofrece una respuesta poderosa al enigma de la menopausia, pero probablemente no constituye toda la respuesta. Como hemos podido ver, explica por qué una mujer podía seguir aumentando su éxito evolutivo después de dejar de tener hijos: alimentando a sus nietos, protegiéndolos, transmitiendo conocimientos, ayudando a sus hijas y permitiendo que una familia sostuviera a más descendientes de los que una madre habría podido criar por sí sola. La supervivencia posreproductiva dejaba así de ser un periodo biológicamente vacío, y aparentemente sin un propósito claro dentro de nuestras sociedades, para convertirse en una nueva forma de inversión familiar.
Sin embargo, demostrar que las abuelas pueden ser beneficiosas no equivale a demostrar que la menopausia evolucionó exclusivamente por ellas. El agotamiento de la reserva ovárica, la pérdida progresiva de calidad de los ovocitos, el aumento de los riesgos asociados al embarazo tardío y la prolongación de la longevidad humana pudieron proporcionar ese sustrato inicial. Sobre él habrían actuado distintas presiones selectivas.
La hipótesis de la madre explica que, llegada cierta edad, proteger a los hijos ya nacidos podía ser más rentable que arriesgar la propia vida en una nueva gestación. La hipótesis del conflicto reproductivo plantea que el cese de la reproducción reducía la competencia entre madres e hijas que comenzaban a reproducirse simultáneamente. La hipótesis de la abuela añade que una mujer todavía sana podía trasladar su esfuerzo hacia los nietos. A ellas se suman la inversión paterna, la ayuda de hermanos mayores, el conocimiento acumulado por los ancianos y las transferencias de alimentos y cuidados dentro de grupos humanos cooperativos.
La menopausia podría haber surgido, por tanto, no como la consecuencia de una única ventaja, sino como parte de una estrategia vital integrada. Una estrategia en la que dejar de producir nuevos descendientes permitía proteger la inversión realizada, reducir conflictos entre generaciones y convertir a los adultos mayores en una fuente de recursos, experiencia y estabilidad para el grupo.
La evolución no “diseñó” la menopausia con un propósito consciente, ni decidió que las mujeres debían convertirse en abuelas. Pero pudo favorecer, generación tras generación, aquellos linajes en los que sobrevivir después de la fertilidad seguía proporcionando más beneficios que costes. Allí donde un nuevo embarazo se volvía menos probable, más peligroso y energéticamente más exigente, la ayuda a los descendientes existentes podría convertirse en una vía alternativa para continuar transmitiendo los propios genes.
La menopausia no representaría entonces una salida de la reproducción, sino un reposicionamiento dentro de ella, que pasaría de engendrar nuevos descendientes a sostener a quienes ya habían nacido.
Críticas y limitaciones de la hipótesis de la abuela
Aunque la hipótesis de la abuela ofrece una explicación convincente para el valor adaptativo de la vida posreproductiva, no resuelve por completo el origen de la menopausia. Explica bien por qué una mujer que ya no se reproduce puede seguir aumentando su aptitud inclusiva, pero resulta menos clara a la hora de explicar cómo apareció inicialmente el cese reproductivo y por qué la selección habría favorecido esa transición.
Además, gran parte de la evidencia disponible es correlacional. Que la presencia de una abuela se asocie con una mayor supervivencia infantil o con una fecundidad más elevada de sus hijas no demuestra necesariamente que la ayuda de la abuela sea la causa directa. La cercanía geográfica, la riqueza familiar, la salud heredada, la estructura del hogar o el acceso compartido a recursos pueden influir también en esos resultados.
El efecto de la abuela tampoco es universal. Depende de su edad, estado de salud, línea de parentesco, proximidad y contexto social. En algunos estudios, las abuelas maternas resultan beneficiosas, mientras que las paternas muestran efectos más débiles o incluso negativos a edades avanzadas. Esto indica que no constituye perse una ventaja automática, sino una estrategia condicionada por la ecología familiar.
La teoría presenta igualmente dificultades para explicar la longevidad masculina, ya que los hombres pueden vivir durante décadas conservando cierta capacidad reproductiva. También debe tener en cuenta los costes fisiológicos de la menopausia, como la pérdida de actividad ovárica y la reducción de estrógenos, que se asocian con un mayor riesgo de osteoporosis y otras enfermedades.
Por último, la hipótesis presupone que las mujeres mayores tuvieron acceso suficiente a sus descendientes, algo que depende de los patrones de residencia y movilidad. En sociedades patrilocales, por ejemplo, las mujeres podían alejarse de su familia materna tras el matrimonio, reduciendo la posibilidad de recibir ayuda de sus propias madres.
Por ello, la hipótesis de la abuela no debería entenderse como una explicación exclusiva. Probablemente, y como ya se ha indicado, describe uno de varios mecanismos que, junto con la protección de los hijos ya nacidos, la reducción del conflicto reproductivo, los costes de la reproducción tardía, la inversión paterna y la crianza cooperativa, pudieron contribuir a mantener y ampliar la vida femenina después de la fertilidad.

Referencias:
https://pmc.ncbi.nlm.nih.gov/articles/PMC2874731/
https://pmc.ncbi.nlm.nih.gov/articles/PMC4356512/
https://nomadwellnessniseko.com/the-female-acl-why-women-are-more-susceptible-to-acl-tears/
https://pubmed.ncbi.nlm.nih.gov/41667102/
https://www.pnas.org/doi/10.1073/pnas.2022159118
https://www.sciencedirect.com/science/article/pii/S1090513817302817
https://data.unicef.org/topic/child-survival/neonatal-mortality/
https://www.who.int/news-room/fact-sheets/detail/maternal-mortality
https://www.cdc.gov/nchs/data/hestat/hestat113.htm
https://ora.ox.ac.uk/objects/uuid%3A9829763f-44ee-423a-a2b1-721f67af3906
https://pubmed.ncbi.nlm.nih.gov/20874739/
https://arxiv.org/pdf/2601.13442
https://www.science.org/doi/10.1126/science.add5473
https://www.diva-portal.org/smash/get/diva2%3A1454442/FULLTEXT01.pdf




